Päivitys 3.5. 2018
TRIM60 , Kr.4q32.3, RNF 129, RNF 33 (C-IV
SPRYPRY) FLJ35882
RING finger
motiivin, B-box2 domaanin, kaksi CC domaania ja B30.2 domaanin
omaava TRIM
TRIM60/RNF33 . Se
tekee interaktion kinesiini-2 perheenjäseniinin Kif3A ja Kif3B
motorisiin proteiineihin, kun ne ovat heterodimerisoituneena ja
kuljettavat kuormaa pitkin mikrotubulusta.Sen on havaittu tekevän
myös proteiini-DNA-interaktioita. Tällä geenillä on pseudogeenejä
ainakin 6 eri kromosomissa (mm. Y kromosomissa, jossa esiintyy trim
vain pseudogeeneineä).
Proteiiniksi asti
koodautuvaa geeniä TRIM60 eeniä esiintyy kahdessa kudoksessa:
testiksessä ja plasentassa (istukassa).
-
- RNF33; RNF129 Summary The protein encoded by this gene
contains a RING finger domain, a motif present in a variety of
functionally distinct proteins and known to be involved in
protein-protein and protein-DNA interactions. Pseudogenes of this
gene are located on more than six chromosomes including chromosome
4. Multiple alternatively spliced variants, encoding the same
protein, have been identified. [provided by RefSeq, Jan 2013]
Expression Low expression observed in reference dataset See
more
Rakenne:
SPRY_PRY_TRIM60;
PRY/SPRY domain of tripartite motif-binding protein 60 (TRIM60) also
known as RING finger protein 33 (RNF33)
zf-B_box; B-box zinc
finger
HMMR_N; Hyaluronan
mediated motility receptor N-terminal
RING-HC_TRIM60_like_C-IV;
RING finger, HC subclass, found in tripartite motif-containing
proteins TRIM60, TRIM61 and similar proteins
DWNN; DWNN domain
PubMed: tripartite motif-containing protein 60
[Homo sapiens]
NCBI Reference
Sequence: NP_001244954.1
LOCUS NP_001244954 471 aa linear PRI 03-OCT-2017
DEFINITION tripartite motif-containing protein 60 [Homo sapiens].
ACCESSION NP_001244954
VERSION NP_001244954.1
DBSOURCE REFSEQ: accession NM_001258025.1
KEYWORDS RefSeq.
SOURCE Homo sapiens (human)
ORGANISM Homo sapiens
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi;
Mammalia; Eutheria; Euarchontoglires; Primates; Haplorrhini;
Catarrhini; Hominidae; Homo.
REFERENCE 1 (residues 1 to 471)
AUTHORS Trynka G, Zhernakova A, Romanos J, Franke L, Hunt KA, Turner G,
Bruinenberg M, Heap GA, Platteel M, Ryan AW, de Kovel C, Holmes GK,
Howdle PD, Walters JR, Sanders DS, Mulder CJ, Mearin ML, Verbeek
WH, Trimble V, Stevens FM, Kelleher D, Barisani D, Bardella MT,
McManus R, van Heel DA and Wijmenga C.
TITLE Coeliac disease-associated risk variants in TNFAIP3 and REL
implicate altered NF-kappaB signalling
JOURNAL Gut 58 (8), 1078-1083 (2009)
PUBMED 19240061
REMARK GeneRIF: Observational study of gene-disease association. (HuGE
Navigator)
REFERENCE 2 (residues 1 to 471)
AUTHORS Saurin AJ, Borden KL, Boddy MN and Freemont PS.
TITLE Does this have a familiar RING?
JOURNAL Trends Biochem. Sci. 21 (6), 208-214 (1996)
PUBMED 8744354
COMMENT REVIEWED REFSEQ: This record has been curated by NCBI staff. The
reference sequence was derived from AC106872.5 and BC100986.2.
Summary: The protein encoded by this gene contains a RING finger
domain, a motif present in a variety of functionally distinct
proteins and known to be involved in protein-protein and
protein-DNA interactions. Pseudogenes of this gene are located on
more than six chromosomes including chromosome 4. Multiple
alternatively spliced variants, encoding the same protein, have
been identified. [provided by RefSeq, Jan 2013].
Transcript Variant: This variant (1) represents the longer
transcript. Both variants 1 and 2 encode the same protein.
Sequence Note: This RefSeq record was created from transcript and
genomic sequence data to make the sequence consistent with the
reference genome assembly. The genomic coordinates used for the
transcript record were based on transcript alignments.
##Evidence-Data-START##
CDS exon combination :: AK093201.1, BX114353.1 [ECO:0000331]
RNAseq introns :: mixed/partial sample support SAMEA1968968,
SAMEA2142853 [ECO:0000350]
##Evidence-Data-END##
FEATURES Location/Qualifiers
source 1..471
/organism="Homo sapiens"
/db_xref="taxon:9606"
/chromosome="4"
/map="4q32.3"
Protein 1..471
/product="tripartite motif-containing protein 60"
/note="ring finger protein 33; ring finger protein 129"
/calculated_mol_wt=54983
Region 13..59
/region_name="RING-HC_TRIM60_like_C-IV"
/note="RING finger, HC subclass, found in tripartite
motif-containing proteins TRIM60, TRIM61 and similar
proteins; cd16607"
/db_xref="CDD:319521"
Region <16 ..105="" region_name="<b">"DWNN"
/note="DWNN domain; cl26688"
/db_xref="CDD:
331509"
Region 16..56
/region_name=
"RING-HC finger (C3HC4-type)"
/note="RING-HC finger (C3HC4-type) [structural motif]"
/db_xref="CDD:
319521"
Site order(16,19,31,33,36,39,53,56)
/site_type="other"
/note="Zn binding site [ion binding]"
/db_xref="CDD:
319521"
Region 92..133
/region_name="
zf-B_box"
/note="B-box zinc finger; pfam00643"
/db_xref="CDD:
306989"
Site order(97,100,119,125)
/site_type="other"
/note="Zn2+ binding site [ion binding]"
/db_xref="CDD:
237988"
Region <139 ..260="" b="">region_name="HMMR_N"
/note
="Hyaluronan mediated motility receptor N-terminal;
pfam15905"
/db_xref="CDD:
318177"
Region 286..465
/region_name
="SPRY_PRY_TRIM60"
/note="PRY/SPRY domain of tripartite motif-binding protein
60 (TRIM60) also known as RING finger protein 33 (RNF33);
cd15828"
/db_xref="CDD:
294000"
CDS 1..471
/gene="TRIM60"
/gene_synonym="RNF129; RNF33"
/coded_by="NM_001258025.1:345..1760"
/db_xref="CCDS:
CCDS3808.1"
/db_xref="GeneID:
166655"
/db_xref="HGNC:
HGNC:21162"
ORIGIN
1 mefvtalvnl qeesscpicl eylkdpvtin cghnfcrscl svswkdlddt fpcpvcrfcf
61 pyksfrrnpq lrnlteiakq lqirrskrkr qkenamcekh nqfltlfcvk dleilctqcs
121 fstkhqkhyi cpikkaasyh reilegslep lrnniervek viilqgsksv elkkkveykr
181 eeinsefeqi rlflqneqem ilrqiqdeem nilaklnenl velsdyvst
l khllrevegk
241 svqsnlellt qaksmhhkyq nlkcpelfsf rltkygfslp pqysg
ldrii kpfqvdvild
301
lntahpqllv sedrkavrye rkkrnicydp rrfyvcpavl gsqrfssgrh ywevevgnkp
361
kwilgvcqdc llrnwqdqps vlggfwaigr ymksgyvasg pkttqllpvv kpskigifld
421
yelgdlsfyn mndrsilytf ndcfteavwp yfytgtdsep lkicsvsdse r
//
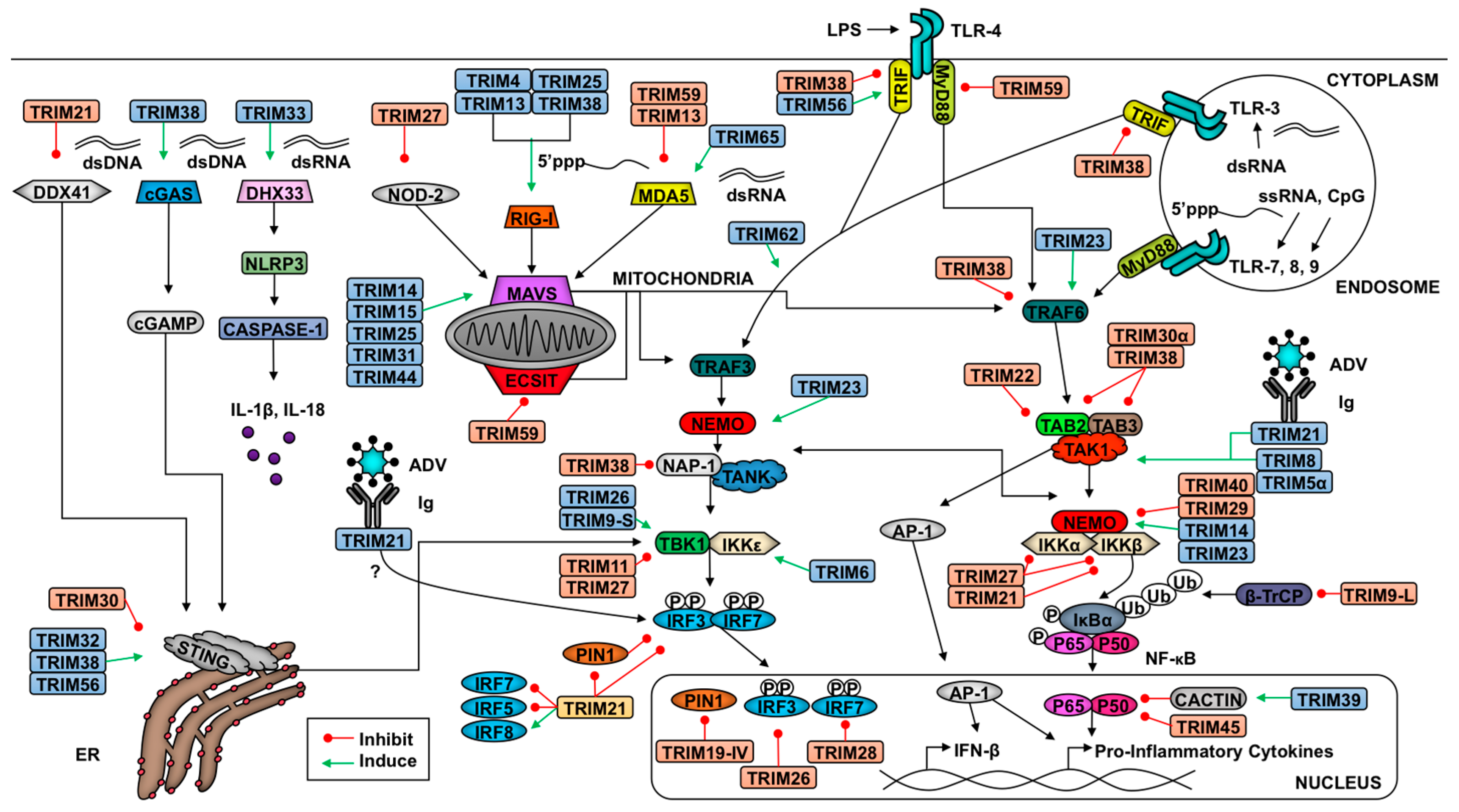
Artikkeleita PubMed . ”TRIM60 PROTEIN” (2)
TRIM60 assosioituu kinesiineihin
RNF33/TRIM60 geeni
on ajallisesti transkriboituna hiien alkiossa
preimplantaatiovaiheessa ennea kuin geeni blastokystivaiheessa
hiljentyy, mutta aikuisella koehiireellä geeni jälleen
transkriboituu testiksessä. Tässä artikkelin tutkimuksessa
tiedemiehet selvittivät TRIM60:n biologista funktiota ja niitä
proteiineja ,jotka assosioituvat TRIM60-proteiiniin. He selvittivät
interaktiodomeenit ja havaitsivat interaktion tekevät
moottoriproteiinit, heterodimeerinä esiintyvät proteiinit KIF3A ja
KIF3B kinesiini-2- perheestä, joiden tiedetään siirtävän
kuormaa pitkin mikrotubulusta. TRIM60-proteiinin RING ja Bbox sekä
PRYSPRY (B30.2) tekivät interaktion KIF3A-KIF2B-heterodimeerin
C-terminaaliseen päätyyn ja heterodimeerin moottoripääty oli
vapaana ja valmiina designtavaroiden kuljetukseen ja
liikkumiseen pitkin mikrotubulusta . Todennäköisesti TRIM60 myös
teki interaktion KIF3A-KIF3B heterodimeerin kanssa kinesiiniin
assosioituvasta soviteproteiinista 3 (KAP3 adaptorista )
riippumattakin Tässä työssä osoitetaan ensimmäsitä kertaa ,
että TRIM60 (RNF33) tekee interaktion kinesiinin molekulaariseen
moottoriin vaikuttaen osaltaan spesifisten kuormien kinesiinistä
riippuvaan mobilisaatioon hiiren testiksen mikrotubuluksissa .
-
The
Rnf33/Trim60 gene is temporally transcribed in the preimplantation
embryo before being silenced at the blastocyst stage but Rnf33
expression is detected in adult testis of the mouse. The putative
RNF33 protein is a tripartite motif (TRIM)/RBCC protein composed of
a typical RING zinc finger, a B-box 2, two α-helical coiled-coil
segments, and a B30.2 domain. As a first step towards the
elucidation of the biologic function of RNF33, we aimed in this
study to elucidate proteins that associate with RNF33.
RNF33-interacting proteins were first derived by the yeast
two-hybrid system followed by co-immunoprecipitation assays.
Interacting domains were determined by deletion mapping in genetic
and biochemical analyzes. RNF33 was shown to interact with the
kinesin-2 family members 3A (KIF3A) and 3B (KIF3B) motor proteins in
the heterodimeric form known to transport cargos along the
microtubule. Domain mapping showed that the RB and B30.2 domains of
RNF33 interacted with the respective carboxyl non-motor domains of
KIF3A and KIF3B. Since RNF33 interacted with the carboxyl-terminal
tail of the KIF3A-KIF3B heterodimer, the motor head section of
KIF3A-KIF3B was free and available for association with designated
cargo(s) and movement along the microtubule. Data also suggest that
RNF33 most likely interacted with KIF3A-KIF3B independent of the
adaptor kinesin-associated protein KAP3. This study is a first
demonstration of a TRIM protein, namely RNF33, that interacts with
the kinesin molecular motors possibly contributing to
kinesin-dependent mobilization of specific cargo(s) along the
microtubule in the testis of the mouse. PMID: 21909995
Similar
articles
-
Select item 212052142.
NF-kB:n säätelemä
TRIM60- ilmenemä erityiseti Sertolin soluissa viittaa siihen, että
TRIM60:lla on ilmeistä osutta spermatogeneesissä kuten edellä
mainittiin osallistumisesta kinesiini-proteiinien suorittamaan
mikrotubuluksia myötäilevään kuljetukseen.
-
Hence,
demonstration of NF-κB-regulated Rnf33 expression in testicular
cells, particularly in Sertoli cells, implicates functional
involvement of the putative RNF33 protein in spermatogenesis through
association of the RNF33 protein with the microtubule via
interaction with kinesin motor proteins, as previously demonstrated
[Huang et al., submitted]. PMID: 21205214
Free
Article Similar
articles
Related
articles in PubMed TRIM60 GENE
-



{kind=link}