Ensiksi näin maininnan proteiinin bromodomeenista TIRMproteiinisa TRIM28 ( KAP1).
Toiseksi siitä mainittiin eräässä väitöstyössä, onkologian alueella. ja siitä kirjoitan tässä enemmän. kolamnneksio taas näin täsä domeenista uudessa immuniteettia kåäsitelevässä kirjassa kirjastossa maintiavan BET bromodomeenista parilla lauseella. Nyt siten huomaan SARS2 viruksen interaktioproteiinien jouksosa kaksi bromodomeenin omaavaa proteiinia BRD2 ja BRD4.
Katson niistäkin geeneistä tiedet tämän teesikommentin jälkeen. Löytyy aika tuore artikkeli ja otan sen seuraavan otsikon alle.
Vuonna 2016 oli väitöstilaisuus ahlgrenskan akatemiassa aiheesta "Targeting Myc-driven tumours BETing on ATR. " Yleensä en niin ensisijaisesti mennyt onkologisia aiheita kuutnelemaan, muta tässä oli tuo MYC, josta yritin saada käsitystä.
Tässä sitten esiintyi se BET bromodomeeni ja HDAC inhibiittorit. (Sirtuiinit olenkin katsonut läpi ja tehnyt niistä muistiinpanpot )
Asetan teesin tiedot tähän: http://hdl.handle.net/2077/41548
Osatyöt:
Bhadury J, Nilsson LM, Muralidharan SV, Green LC, Li Z, Gesner EM,
Hansen HC, Keller UB, McLure KG, Nilsson JA.
BET and HDAC inhibitors
induce similar genes and biological effects and synergize to kill in
Myc-induced murine lymphoma. Proceedings of the National Academy of
Sciences. 2014 Jul 1;111(26):E2721-30
VISA ARTIKEL
II.
Muralidharan SV, Bhadury J, Nilsson LM, Green LC, McLure KG, Nilsson
JA.
BET bromodomain inhibitors synergize with ATR inhibitors to induce
DNA damage, apoptosis, senescence-associated secretory pathway and ER
stress in Myc-induced lymphoma cells. Oncogene. 2016 Jan 25
VISA ARTIKEL
III.“Synergistic
enhancement of apoptosis in melanoma by ATR & BET bromodomain
inhibitors”. Somsundar Veppil Muralidharan, Berglind Einarsdottir,
Mattias Lindberg, Joydeep Bhadury, Eric Campeau, Roger Olofsson Bagge,
Ulrika Stierner, Lars Ny, Lisa M. Nilsson and Jonas A. Nilsson
(MANUSCRIPT).
Cancer arises from loss of function of tumour suppressors and/or
gain of function mutations in proto-oncogenes that disrupt the delicate
balance required for homeostatic cell division, resulting in
uncontrolled cell proliferation.
Oncogenic transformation of
multifaceted proto-oncogene - transcription factor - MYC can give rise
to cancers and it is found to be deregulated in more than 70% of the
tumours.
Targeting MYC directly or identifying the Achille’s heel of
MYC-driven tumours is thus a promising
therapeutic approach to treat these tumours.
This thesis investigates
and demonstrates novel therapeutic approaches against MYC-driven
tumours.
In the first publication (Bhadury et al, 2014), we characterize a novel
and orally bio-available BET bromodomain inhibitor (BETi) RVX2135.
We
also identified BET bromodomain proteins as a valuable therapeutic
target against MYC driven tumours in vitro and in vivo.
Gene expression
profiling to identify these transcriptional changes enabled us to
identify subset of genes that are commonly altered by both BETi and
HDACi.
This study also demonstrates that HDACi and BETi can synergize to
hinder Myc-induced lymphoma progression.
The second publication (Muralidharan et al, 2016) in this thesis
investigates the role of BET proteins in regulating cell cycle and
replication.
BETi disable the entry of cells into S-phase of cell-cycle,
hamper DNA synthesis and cause DNA damage.
A pharmacogenetic screen
identified BET inhibitors to synergize with inhibition of PI3K/mTOR
family of proteins, to which ATR, an upstream kinase of DDR pathway
belongs.
Further studies revealed that the thus identified PI3K/mTOR
inhibitors indeed affect ATR-Chkl DDR pathway leading to the discovery
of a strong synergy between BETi and ATRi in apoptosing Myc driven
tumours in vitro, and in vivo and (by) it induces SASP and ER stress.
The third study translates the above findings into the field of
melanoma, a form of skin cancer. We validate the BETi-ATRi synergy in
cell lines in vitro and in Patient Derived Xenografts (PDX) in vivo.
Using B16F10 in vivo syngenic transplant melanoma model, we also
demonstrated that this combination therapy can be safely combined with
Immune Therapy, the front line treatment against melanoma in clinic
today.
Taken together, this thesis puts forth a multifaceted approach to treat
cancer. It thoroughly describes the effects of BETi and ATRi on cancer
cells and how they can be combined to enhance the therapeutic efficacy

Visar inlägg med etikett c-Myc. Visa alla inlägg
Visar inlägg med etikett c-Myc. Visa alla inlägg
torsdag 9 juli 2020
onsdag 8 oktober 2014
c-Myc, BMP signalointi, SMAD-1,-5,-8

FIGURE 7
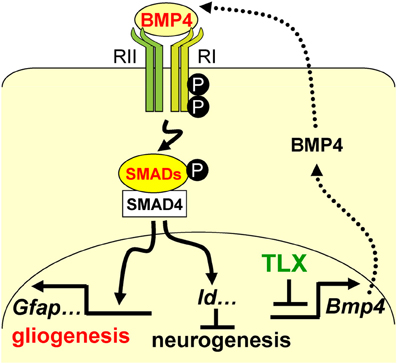 Figure 7. A schematic diagram illustrating the interplay
between TLX and the BMP-SMAD signaling pathway in the regulation of
neurogenesis and astrogenesis. TLX directly modulates the
expression of the BMP ligands, which bind and activate the type I (RI)
and type II (RII) receptors. These events result in phosphorylation of
regulatory SMADs and their dimerization with the common cofactor SMAD4.
The SMAD complex acts as a transcriptional activator to induce the
expression of downstream targets, which promote astrogenesis and inhibit
neurogenesis.
Figure 7. A schematic diagram illustrating the interplay
between TLX and the BMP-SMAD signaling pathway in the regulation of
neurogenesis and astrogenesis. TLX directly modulates the
expression of the BMP ligands, which bind and activate the type I (RI)
and type II (RII) receptors. These events result in phosphorylation of
regulatory SMADs and their dimerization with the common cofactor SMAD4.
The SMAD complex acts as a transcriptional activator to induce the
expression of downstream targets, which promote astrogenesis and inhibit
neurogenesis.c-Myc geeni ja sen funktioista. Merkitys onkogeeninä.
http://en.wikipedia.org/wiki/Myc
Myc (c-Myc) is a regulator gene that codes for a transcription factor. The protein encoded by this gene is a multifunctional, nuclear phosphoprotein that plays a role in cell cycle progression, apoptosis and cellular transformation.
Myc- vikatoimintaa on havaittu myös kohdunkaulan syövässä, paksunsuolen syövässä, rinta-, keuhko- ja mahasyövässä. Myc onkin lupaava antisyöpälääkkeiten kohde.
A mutated version of Myc is found in many cancers, which causes Myc to be constitutively (persistently) expressed. This leads to the unregulated expression of many genes, some of which are involved in cell proliferation, and results in the formation of cancer. A common human translocation involving Myc is critical to the development of most cases of Burkitt's Lymphoma Malfunctions in Myc have also been found in carcinoma of the cervix, colon, breast, lung and stomach. Myc is thus viewed as a promising target for anti-cancer drugs.
In the human genome, Myc is located on chromosome 8 and is believed to regulate expression of 15% of all genes[4] through binding on Enhancer Box sequences (E-boxes) and recruiting histone acetyltransferases (HATs). This means that in addition to its role as a classical transcription factor, Myc also functions to regulate global chromatin structure by regulating histone acetylation both in gene-rich regions and at sites far from any known gene.
Myc protein belongs to Myc family of transcription factors, which also includes N-Myc and L-Myc genes. Myc family of transcription factors contain bHLH/LZ (basic Helix-Loop-Helix Leucine Zipper) domain. Myc protein, through its bHLH domain can bind to DNA, while the leucine zipper domain allows the dimerization with its partner Max, another bHLH transcription factor.
Myc mRNA contains an IRES (internal ribosome entry site) that allows the RNA to be translated into protein when 5' cap-dependent translation is inhibited, such as during viral infection.
Myc is activated upon various mitogenic signals such as Wnt, Shh and EGF (via the MAPK/ERK pathway). By modifying the expression of its target genes, Myc activation results in numerous biological effects. The first to be discovered was its capability to drive cell proliferation (upregulates cyclins, downregulates p21), but it also plays a very important role in regulating cell growth (upregulates ribosomal RNA and proteins), apoptosis (downregulates Bcl-2), differentiation, and stem cell self-renewal. Myc is a very strong proto-oncogene and it is very often found to be upregulated in many types of cancers. Myc overexpression stimulates gene amplification,[7] presumably through DNA over-replication.
There have been several studies that have clearly indicated Myc's role in cell competition.
A major effect of Myc is B cell proliferation.
c-Myc induces AEG-1 or MTDH gene expression and in turn itself requires AEG-1 oncogene for its expression.
Alla olevissa kohdissa on tärkeää suomennettavaa vielä:

- Myc ( c-Myc) on säätelijägeeni, joka koodaa yhtä transkriptiotekijää . Tämän geenin koodaama proteiini on multifunktionaalinen nukleaarinen fosfoproteiini, jolla on tehtävää solusyklin progressiossa, apoptoosissa ja solun transformaatiossa.
Myc (c-Myc) is a regulator gene that codes for a transcription factor. The protein encoded by this gene is a multifunctional, nuclear phosphoprotein that plays a role in cell cycle progression, apoptosis and cellular transformation.
- Monessa syövässä on havaittu sellainen mutatoitunut muoto c-Myc-geeniä. että se on jäänyt pinttyneesti päälle, konstitutiivisesti ilmeneväksi. Tästä seuraa monien geenien säätelemätöntä ilmenemistä ja moni niistä osallistuu solun proliferaatioon ja johtaa sen takia erilaisiin syöpämuodostumiin.
Myc- vikatoimintaa on havaittu myös kohdunkaulan syövässä, paksunsuolen syövässä, rinta-, keuhko- ja mahasyövässä. Myc onkin lupaava antisyöpälääkkeiten kohde.
A mutated version of Myc is found in many cancers, which causes Myc to be constitutively (persistently) expressed. This leads to the unregulated expression of many genes, some of which are involved in cell proliferation, and results in the formation of cancer. A common human translocation involving Myc is critical to the development of most cases of Burkitt's Lymphoma Malfunctions in Myc have also been found in carcinoma of the cervix, colon, breast, lung and stomach. Myc is thus viewed as a promising target for anti-cancer drugs.
- Ihmisen genomissa Myc sijaitsee kromosomissa 8 ja arvellaan sen säätelevän 15%: kaikkien geenien ilmenemästä siten, että se sitoutuu Enchancer Box- sekvensseihin ( E- Box), jotka ovat kiihdyttäjäjaksoja ja rekrytoi histoniasetyylitransferaaseja (HAT entsyymejä) . Tämä merkitsee, että se on klassinen transkriptiotekijä ja sen lisäksi vielä toimii globaalisti ( koko genomin) kromatiinistruktuurin säätelyssä säätelemällä histoniasetylaatioita sekä geenipitoisilla alueilla että niillä kohdilla, jotka sijaitsevat kaukana kaikista tunnetuista geeneistä.
In the human genome, Myc is located on chromosome 8 and is believed to regulate expression of 15% of all genes[4] through binding on Enhancer Box sequences (E-boxes) and recruiting histone acetyltransferases (HATs). This means that in addition to its role as a classical transcription factor, Myc also functions to regulate global chromatin structure by regulating histone acetylation both in gene-rich regions and at sites far from any known gene.
- 1. Miten Myc keksittiin? Tämä tapahtui Burkitin lymfoomapotilaasta. Syöäpäsoluissa oli kromosomaalinen translokaatio ja siinä usein oli kromosomi 8 osallisena. kun kloonattiin fuusiokromosomin katkoskohta paljastui geeni,m joka oli samanlainen kuin myelosytomatoosin virusperäinen onkogeeni v-Myc. Niimpä uusi soluperäinen (cellular) geeni nimettiin c- Myc- nimellä.
- 2. Minkälinen on Myc rakenne? Myc-proteiini kuuluu Myc-transkiptiotekijöiden perheeseen, johon kuuluu myös N-Myc ja L-Myc geenit Myc-perheessä on bHLH(LZ ( baasinen Helix-silmukka- Helix Leusiini Zipper- domaani . Myc-proteiini voi tuolla bHLH domaanilla sitoutua DNA:han kun leusiini-zipper- domaani taas antaa dimerisoitumismahdollisuuden Max- partnerin kanssa- se on toinen bHLH transkriptiotekijä. . Myc- mRNA sisältää IRES ( internal ribosome entry site) kohdan , joka sallii RNA:n translaation proteiiniksi, jos 5prim cap- riippuvainen translaatio on estyneenä, kuten virusinfektiossa.
Myc protein belongs to Myc family of transcription factors, which also includes N-Myc and L-Myc genes. Myc family of transcription factors contain bHLH/LZ (basic Helix-Loop-Helix Leucine Zipper) domain. Myc protein, through its bHLH domain can bind to DNA, while the leucine zipper domain allows the dimerization with its partner Max, another bHLH transcription factor.
Myc mRNA contains an IRES (internal ribosome entry site) that allows the RNA to be translated into protein when 5' cap-dependent translation is inhibited, such as during viral infection.
- 3. Mitkä ovat Myc toiminnot? Kuten jo yllä mainittiin myc proteiini on transkriptiotekijä, joka aktivoi monen geenin ilmenemän sitoutumalla konsensusjaksoihin( E-box9 ja rekrytoimalla histoniasetyylitransferaseja. Muta se voi toimia myös transkirptionaalisena vaimentajana. Jos se sitoutuu miz-q transkirptiotekijöäön ja preijaa paikaltaan p300 koaktivaattorin, se estää Miz-1 kohdegeenien ilmenemistä. Lisäksi myc omaa suoraa osuutta DNA-replikaation kontrollissa. Moni mitogeeni signaali on Myc- aktivaation taustana, kuten Wnt, Shh ja EGF ( MAPK/ERK teitse) . Myc- aktivaatio johtaa lukuisiin biologisiin vaikutuksiin, koska se ystyy modifioimaan kohdek´geeniensä ilmenemistä. Ensinnäkin havaittiin sen kyky panna liikkeelle soluproliferaatio ( säätää ylös sykliineitä, vaimentaa p21). muta sen lisäksi sillä on hyvin tärkeää osaa solukasvun säätelyssä ( se säätää ylös ribosomaalista RNA.ta ja proteiineja) , se vaikutaa apoptoosia ( säätää alas Bcl-2), solujen erikoistumista ja kantasolujen itsenäistä uudistumista. myc on hyvin vahva proto-onmkogeeni ja hyvin usein sitä havaitaan monen tyyppisissä syövissä ylössäätyneessä tilassa. Jos Myc ilmenee liiallisena, stimuloituu geenin amplifikaatio, oletettavsti DNA:n liikamääräisenä replikoitumisena
- On useita tutkimuksia, jossa on viitettä siitä, että Myc omaa osuutta solujen keskeisessä kilpailussa . Eräs sen suuri vaikutus ilmenee B-solujen proliferaatiossa
- c-Myc indusoi AEG-1 tai MTDH geenin ilmenemän ja puolestaan itse vaatii AEG-1 onkogeeniä omaan ilmenemiseensä.
Myc is activated upon various mitogenic signals such as Wnt, Shh and EGF (via the MAPK/ERK pathway). By modifying the expression of its target genes, Myc activation results in numerous biological effects. The first to be discovered was its capability to drive cell proliferation (upregulates cyclins, downregulates p21), but it also plays a very important role in regulating cell growth (upregulates ribosomal RNA and proteins), apoptosis (downregulates Bcl-2), differentiation, and stem cell self-renewal. Myc is a very strong proto-oncogene and it is very often found to be upregulated in many types of cancers. Myc overexpression stimulates gene amplification,[7] presumably through DNA over-replication.
There have been several studies that have clearly indicated Myc's role in cell competition.
A major effect of Myc is B cell proliferation.
c-Myc induces AEG-1 or MTDH gene expression and in turn itself requires AEG-1 oncogene for its expression.
Alla olevissa kohdissa on tärkeää suomennettavaa vielä:
- 4 Myc-nick
- 5 Clinical significance
- 6 Animal Models
- 7 Interactions
- (Tässä mainitaan mm. interaktiot SMAD2 ja SMAD3 kanssa)
- 8 See also
- 9 References
- 10 Further reading
- 11 External links
Prenumerera på:
Kommentarer (Atom)