
25.5. 2020 Kertaan SMAD interaktomia ja signalointia, koska löysin maininnan että SARS2 virusproteiini N saattaa tehdä interaktion SMAD3 proteiinin kanssa. Myös SNIP1 on sen interaktioproteiineja: (SNIP1 , SMAD nuclear interacting protein 1, (1p34.3)
Lisäksi SMAD3 osallistunee ihmisen RNA alueeseen, joka nyt alkaa herättää huomota myöls SARS2 viruksen kapasiteettialueena. on myös lisätietoa SMAD interaktomsta tästä lähteestä: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5104937/.
........
Päivitys 22.5. 2019
Lisäksi SMAD3 osallistunee ihmisen RNA alueeseen, joka nyt alkaa herättää huomota myöls SARS2 viruksen kapasiteettialueena. on myös lisätietoa SMAD interaktomsta tästä lähteestä: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5104937/.
........
Päivitys 22.5. 2019
SMAD signalointi
https://www.qiagen.com/sg/shop/genes-and-pathways/pathway-details/?pwid=412
https://www.qiagen.com/geneglobe/static/images/Pathways/SMAD%20Signaling%20Network.jpg
https://www.qiagen.com/geneglobe/static/images/Pathways/SMAD%20Signaling%20Network.jpg
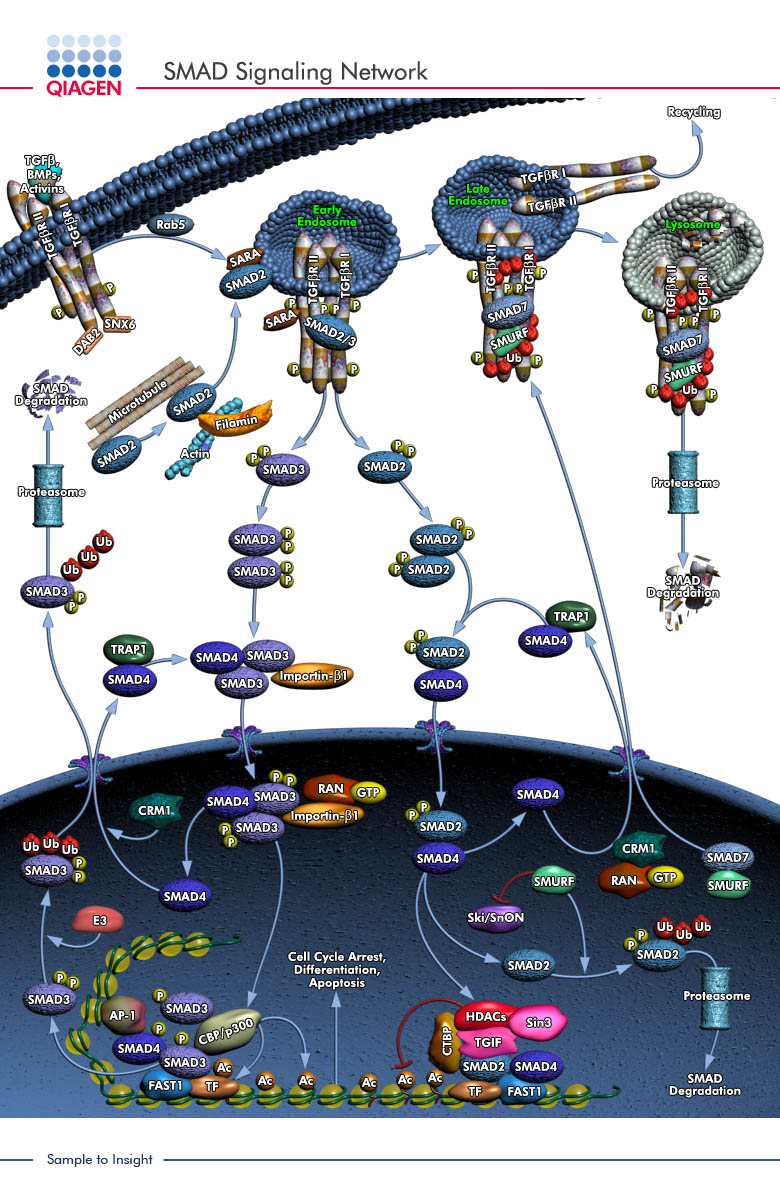{kind=link}
SMAD- signaaliverkostosta
( Käännöstä
artikkelista netistä: Kirjoittajan nimeä en havaitse tekstissä)
Verisuoniston sisällä
esiintyvä erittyneiden polypeptidien TGF- beeta-superperhe
on kasvutekijöitä, jotka ovat tärkeitä vaikuttaja
patofysiologisissa prosesseissa. Näihin prosesseihin kuuluu
angiogeneesi, verisuoniston uudelleenmuovautuminen, aterogeneesi
ja osallistuminen soluvasteiden säätelyyn ( kasvun,
proliferaation, erilaistumisen, migroitumisen, adhesoitumisen, solun
elossapysymisen ja kehityksellisen solukohtalon säätelyyn).
TGF-beetan
ohella Aktiviinit ja BMP- proteiinit
kuuluvat mainittuun superperheeseen. Nämä tekijät
signaloivat heteromeeriksi koostuneen
reseptorikompleksin kautta.
TGF-beta-reseptorit ovat
seriini/treoniini(S/T)kinaasityyppiset I ja II.
TGF-beeta-reseptorisignaloinnin
alavirrassa on SMAD- proteiinien signaaliverkosto.
TGF-beta= Transforming
Growth factor beta, kasvua muuntava kasvutekijä
BMP = Bone
Morphogenetic Protein, luun
morfogeneettinen proteiini
SMAD = (family of
transductin pathway; signaalia johtava proteiiniperhe);
(SMA and MAD related family).
Nimi SMAD tulee
fylogeneettisesti ensimmäiseksi löytyneestä SMAD-proteiinista
(banaanikärpäsestä Mothers Against Decapentaplegic)
Näillä SMAD-proteiineilla
on ratkaiseva osuus monisoluisen organismin solukohtaloiden
päättämisessä.
-
SMAD Signaling Network.
-
Within the vasculature, TGF-Beta (Transforming Growth Factor-Beta) superfamily of secreted polypeptide growth factors play an important role in a variety of pathophysiologic processes, including angiogenesis, vascular remodeling, atherogenesis and in regulating cellular responses such as growth, proliferation, differentiation, migration, adhesion, survival, and specification of developmental fate. Apart from TGF-Beta, the superfamily also includes the Activins and the BMPs (Bone Morphogenetic Proteins). These factors signal through heteromeric complexes of Type-II and Type-I serine-threonine kinase receptors, which activate the downstream SMAD (Sma and Mad Related Family) signal transduction pathway (Ref.1, 2 & 3).
Rakenteen ja funktionaalisten tehtävien perusteellä
SMAD-perheen jäsenet (MADH) jaetaan lämminverisillä ainakin
kolmeen laajaan luokkaan.
(1) Co-SMAD, kooperatiiviset SMAD- molekyylit, joita on SMAD4/DPC4 ja SMAD10 ja ne osallistuvat TGF-beeta-perheen jäsenten signalointiin.
(2) R- SMAD prroteiinit. Nämä ovat reseptorin säätelemiä SMAD- proteiineja ja niihin kuuluu SMAD1, SMAD2, SMAD3, SMAD5 ja SMAD8/9. Niistä jokaisella on oma spesifinen signaalitiensä.
(3) I-SMAD-proteiinit, inhibitoriset, antagonistiset SMAD6 ja SMAD7- proteiinit. Niiden säätely signaalitiessä on negatiivista.
(1) Co-SMAD, kooperatiiviset SMAD- molekyylit, joita on SMAD4/DPC4 ja SMAD10 ja ne osallistuvat TGF-beeta-perheen jäsenten signalointiin.
(2) R- SMAD prroteiinit. Nämä ovat reseptorin säätelemiä SMAD- proteiineja ja niihin kuuluu SMAD1, SMAD2, SMAD3, SMAD5 ja SMAD8/9. Niistä jokaisella on oma spesifinen signaalitiensä.
(3) I-SMAD-proteiinit, inhibitoriset, antagonistiset SMAD6 ja SMAD7- proteiinit. Niiden säätely signaalitiessä on negatiivista.
-
Based on their structures and known functional roles, the mammalian SMAD family members (Mad-homologues, MADH) fall into at least three broad classes:
(i) the Co-SMADs (Co-mediator SMADs), SMAD4/DPC4
and SMAD10, participate in signaling by diverse TGF-Beta
family members;
(ii) the R-SMADs (Receptor-regulated SMADs), including SMAD1, SMAD2, SMAD3, SMAD5, and SMAD8, which are each involved in a specific signaling pathways; and
(iii) the antagonistic SMADs, including SMAD6 and SMAD7, which negatively regulate these pathways (Ref.1).
TGF-beeta reseptorit ja aktiviinireseptorit fosforyloivat ( eli aktivoivat) SMAD2 ja SMAD3- proteiinit.
BMP-reseptorit fosforyloivat SMAD1-, SMAD5- ja SMAD8-proteiinit.
Jotta erityinen TGF-beetavaste pääsisi tapahtumaan, TGF-beeta- superperheen jäsenen tulee dimerisoituneena asettua korkean affiniteetin reseptoriinsa (Tyyppi II-reseptori) ja I-tyypin reseptori transfosforyloituu. Täten aktivoitunut TGF-reseptorikompleksi alkaa rekrytoida tekijöitä, jotka edistävät signaalin jatkumista solun sisälle (DAB2, SNX6).
Pienet GTPaasit (Rab5) katalysoivat aktivoituneen reseptorin pääsyä varhaiseen endosomiin, jossa ne kohtaavat fosfolipideihin sitoutuneen kuljettajan kuten SARA.
SARA on reseptorin aktivoitumiseen tarvittava SMAD-Ankkuri ja se avustaa SMAD-molekyylien rekrytoimisessa I-tyypin kinaasille. SARA ei tee interaktiota SMAD1- tai SMAD5-proteiinien kanssa.
(ii) the R-SMADs (Receptor-regulated SMADs), including SMAD1, SMAD2, SMAD3, SMAD5, and SMAD8, which are each involved in a specific signaling pathways; and
(iii) the antagonistic SMADs, including SMAD6 and SMAD7, which negatively regulate these pathways (Ref.1).
TGF-beeta reseptorit ja aktiviinireseptorit fosforyloivat ( eli aktivoivat) SMAD2 ja SMAD3- proteiinit.
BMP-reseptorit fosforyloivat SMAD1-, SMAD5- ja SMAD8-proteiinit.
Jotta erityinen TGF-beetavaste pääsisi tapahtumaan, TGF-beeta- superperheen jäsenen tulee dimerisoituneena asettua korkean affiniteetin reseptoriinsa (Tyyppi II-reseptori) ja I-tyypin reseptori transfosforyloituu. Täten aktivoitunut TGF-reseptorikompleksi alkaa rekrytoida tekijöitä, jotka edistävät signaalin jatkumista solun sisälle (DAB2, SNX6).
Pienet GTPaasit (Rab5) katalysoivat aktivoituneen reseptorin pääsyä varhaiseen endosomiin, jossa ne kohtaavat fosfolipideihin sitoutuneen kuljettajan kuten SARA.
SARA on reseptorin aktivoitumiseen tarvittava SMAD-Ankkuri ja se avustaa SMAD-molekyylien rekrytoimisessa I-tyypin kinaasille. SARA ei tee interaktiota SMAD1- tai SMAD5-proteiinien kanssa.
-
TGF-Beta Receptors and Activin Receptors phosphorylate SMAD2 and SMAD3, and BMP receptors phosphorylate SMAD1, SMAD5 and SMAD8. To initiate a particular TGF-Beta response, dimeric ligands of the TGF-Beta superfamily bind with high affinity to Type-II receptor and trans-phosphorylate Type-I receptor serine/threonine kinases on the cell surface. Activated receptors recruit adaptor proteins such as DAB2 (Disabled-2) and SNX6 (Sortin Nexin-6) that positively affect signal transduction (Ref.5). Small GTPases such as Rab5 catalyze movement of activated receptor complexes to early endosomal compartments (Ref.6), where they encounter phospholipid-bound carriers such as SARA (SMAD Anchor for Receptor Activation) that assist in recruitment of the SMADs to the Type-I receptor kinase. SARA does not interact with either SMAD1 or SMAD5 (Ref.7).
Kun R-SMAD- proteiinit SMAD2 ja SMAD3 sitoutuvat
fosforyloituneeseen GS- domeeniin fosfoseriiniä sitovalla
domeenillaan MH2 ( Mad Homology-2), seuraa niiden irtautuminen
reseptorista ja SARAsta. Muut SMAD-proteiinit tunnistavat
MH2-domeenillaan näitä fosfoseriinejä ja seuraa
homo-oligomerisaatio R-SMAD- proteiinien kesken tai
hetero-oligomerisaatio ainutlaatuisen Co-SMAD kanssa (SMAD4/DPC).
-
The binding of R-SMADs, SMAD2 and SMAD3, to the phosphorylated GS domain via their phosphoserine-binding MH2 (Mad Homology-2) domain leads to its rapid dissociation from the receptor and SARA. These phosphoserines are recognized by the MH2 domain of another SMAD leading to homo-oligomerisation of R-SMADs or hetero-oligomerisation with the unique Co-SMAD (SMAD4/DPC4 in mammals).
SMAD4/DCP4 on sytoplasmassa
ankkuroituneena rakenteelliseen proteiiniin TRAP1
(TGF-beeta reseptorityyppi I:een assosioituneeseen proteiiniin 1),
joka edistävästi avustaa R-SMAD/Co-SMAD-oligomerisaatiota.
Fosforyloitunut SMAD3 assosioituu taas Importiini-beeta1-proteiiniin ja kulkeutuu tumaan. Nukleoplasmassa (tumassa) oleva RanGTPaasi vapauttaa SMAD3-kompleksin (SMAD4 palaa sytoplasmaan ja assosioituu TRAP1:een).
Mutta fosforyloitunut (aktiivi) SMAD2 ei taas pysty sitomaan importiineja ja se kulkee autonomisesti tumaan.
Perustilassaan SMAD4 /DPC4 pääsee konstitutiivisesti tumaan, mutta exportiini (CRM1/XPO) kuljettaa sen saman tien takaisin sytoplasmaan ( Shuttling between nucleus and cytoplasma, constitutively).
Mutta jos on tapahtunut TGF-beetastimulaatio , SMAD4/DPC4 saapuu tumaan kompleksina R-SMAD-proteiinin kanssa (R-SMAD/Co-SMAD) ja säätelee geeniexpressiota.
Fosforyloitunut SMAD3 assosioituu taas Importiini-beeta1-proteiiniin ja kulkeutuu tumaan. Nukleoplasmassa (tumassa) oleva RanGTPaasi vapauttaa SMAD3-kompleksin (SMAD4 palaa sytoplasmaan ja assosioituu TRAP1:een).
Mutta fosforyloitunut (aktiivi) SMAD2 ei taas pysty sitomaan importiineja ja se kulkee autonomisesti tumaan.
Perustilassaan SMAD4 /DPC4 pääsee konstitutiivisesti tumaan, mutta exportiini (CRM1/XPO) kuljettaa sen saman tien takaisin sytoplasmaan ( Shuttling between nucleus and cytoplasma, constitutively).
Mutta jos on tapahtunut TGF-beetastimulaatio , SMAD4/DPC4 saapuu tumaan kompleksina R-SMAD-proteiinin kanssa (R-SMAD/Co-SMAD) ja säätelee geeniexpressiota.
-
SMAD4/DPC4 is anchored to the cytoplasm by scaffolding proteins such as TRAP1 (TGF-Beta Receptor Type-I Associated Protein-1), which assist positively in R-SMAD/Co-SMAD oligomerisation (Ref.4). Phosphorylated SMAD3 associates with Importin-Beta1 and is imported to the nucleus. The Ran GTPase catalyses the transport and release of the SMAD3 complex in the nucleoplasm. In contrast, phosphorylated SMAD2 fails to bind to Importins and is autonomously imported to the nucleus. In the ground state, SMAD4/DPC4 enters the nucleus constitutively and is immediately exported back to the cytoplasm by the Exportin CRM1/XPO1. But, after TGF-Beta stimulation, SMAD4/DPC4 enters the nucleus in complex with R-SMADs (R-SMAD/Co-SMAD complexes) and regulates gene expression.
Sekä SMAD3 että SMAD4/DPC4 sitoutuvat
SBE -elementtiin (SMAD:ia sitoviin vaste elementihin) DNA
sekvenssissä. Mutta päinvastoin SMAD2 ei pysty sitoutumaan SBE-
elementtiin; silti se voi osallistua DNA:lle sitoutuneeseen
kompleksiin, jos se on tehnyt interaktion SMAD4/DPC4:n kanssa
ja silloin se aktivoi spesifisten geenien ilmenemää
ko-operatiivisin interaktioin DNA:ta sitovien proteiinien
ohella (mm transkriptiofaktoriperheen winged - helix eli
FOX- perheen jäsenten kanssa) FAST1 ja FAST2 (Forkhead
Activin signal transducers ).
-
Both SMAD3 and SMAD4/DPC4 bind the SBE (SMAD-Binding Elements) to DNA sequences. In contrast, SMAD2 fails to bind to SBEs but it participates in DNA-bound complexes via its interaction with SMAD4/DPC4, and activates expression of specific genes through cooperative interactions with DNA-binding proteins, including members of the winged-helix family of TFs (Transcription Factors), FAST1 and FAST2 (Forkhead Activin Signal Transducers) (Ref.6).
Lisäksi sekä R-SMAD- proteiinit että
Co-SMAD tekevät interaktioita monien yleisten ja kudosspesifisten
transkriptiotekijöiden kanssa MH1- ja MH2-domeeneillaan.
Tumassa olevien SMAD-kompleksien transkriptioaktiivisuutta
moduloi DNA:ta sitova proteiini TGIF (TGF-beetan indusoima
faktori), proto-onkogeenit Ski ja SnoN, ja ne vaikuttavat
SMAD-transkriptioon ko-repressoreina ( myötävaimentajina).
-
In addition, both R-SMADs and the Co-SMAD interact with many general and tissue-specific TFs via their MH1 or MH2 domains. The transcriptional activity of nuclear SMAD complexes within the nucleus is modulated by DNA-binding protein TGIF (TGF-Beta Induced Factor), proto-oncogene Ski and SnoN, which act as SMAD transcriptional co-repressors (Ref. 7,9).
Sellaisetkin transkriptiotekijät, jotka itse eivät
sido DNA:ta , assosioituvat tuman puolella oleviin
SMAD-proteiineihin ja rekrytoivat koaktivaattoreita, kuten
CBP(CREB-binding protein)/p300 (HAT), jotka johtavat
nukleosomaalisten histoneiden asetyloitumiseen, ja/tai
proteiineja jotka ovat assosioituneita
transkriptiotekijöitä ja ratkaisevia transkription
indusoitumisessa.
CREB=
cAMP responsive element binding protein 1 (2q33.3)
p300
(22q13.2) histone acetyltransferase, mediates cAMP gene
regulation by binding to phosphopylated CREB protein)
Vaihtoehtoisesti tumassa olevat SMAD-proteiinit
rekrytoivat ko-repressiivisiä tekijöitä, jotka ovat
histonideasetylaaseihin (HDAC) liittyviä; niitä
ovat CTBP ( C-terminaalisesti sitova proteiini) ja Sin3 (HDAC
alayksikkö), ja nämä johtavat transkription
repressoitumiseen kohdegeeneillä (acetylryhmät irtoavat
histoneista).
SIN3,
HDAC complex subunit sin3a, transcriptional corepressor Sin3a,
WITKOS
CTBP1 ,
BARS, HADDTS (4p16.3) , dimerise CTBP2. Transcription repressor
-
Non-DNA-binding TF also associate with nuclear SMADs and recruit co-activators such as CBP (CREB-Binding Protein)/p300 that lead to acetylation of nucleosomal histones and/or associated TF, which are crucial for transcriptional induction. Alternatively, the nuclear SMADs also recruit co-repressors that associate with HDACs (Histone Deacetylases) such as CTBP (C-Terminal Binding Protein) and Sin3, thus leading to transcriptional repression of target genes.
R-SMAD proteiinit 1, 2 ja 3 saattavat
liikkua itsenäisesti sytoplasmasta tuman puolelle, mutta SMAD4:n
täytyy ensin tehdä kompleksi jonkin näistä
kanssa ennen kuin se pääsee tumaan. Tuman SMAD-proteiinit voivat
osallistua myös ubikitinoitumisreaktioihin, jotka johtaa
signaalitien alassäätymiseen tai muiden
transkriptiotekijöiden hajoamiseen.
Fosforyloitunut tumassa oleva SMAD3 ubikitinoituu E3
ligaasilla Roc1/SCF, kun on tehnyt transkriptiivisen työnsä, ja
sitten kulkeutuu sytoplasmaan proteosomaaliseen silppuroitumiseen.
Sytoplasmiset R- SMAD- proteiinit solun perustilassa
joutuvat SMAD-spesifisen E3-ligaasiperheen substraateiksi.
SMURF-perhe E3 ligaaseja ovat SMAD-proteiineja
ubikitiinilla sääteleviä tekijöitä ja johtavat myös R- SMAD:in
proteosomisilppuriin ja pitävät saatavilla olevaa R-SMAD-
allasta matalapitoisena. Vaihtoehtoisesti tumassa oleva
R-SMAD-SMURF-kompleksi voi hyökätä transkription vaimentajaa SnoN
kohtaan ja säätää vaimentajan alas.
-
Nuclear SMADs also participate in ubiquitination reactions that lead to downregulation of the pathway itself or degradation of other TFs.
Kolmas SMAD proteiiniluokka I-SMAD on
SMAD-inhibiittoreita , kuten SMAD7. SMAD7 estää R-
SMAD-proteiinien rekrytoimista ja niiden fosforyloimista
(aktivoimista) . Lisäksi SMAD7 assosioituu E3 ubikitiiniligaasien
SMURF:ien kanssa ja muodostaa SMAD7-SMURF kompleksin, kun
TGF-beeta-stimulaatio on lähtenyt liikkelle ja se kompleksi
ubikitinoi reseptoreita solupinnalla tai endosomaalisella kalvolla
ja kohdentaa proteiinisilppuriin proteosomeihin tai
lysosomeihin.
Eräs adaptoriproteiini STRAP1 sitoutuu myös
I-tyypin reseptoreihin ja myös SMAD7- proteiiniin ja
vahvistaa SMAD7- proteiinin inhibitorista aktiivisuutta.
-
Third classes of SMADs , the I-SMADs, such as SMAD7 inhibit the recruitment and phosphorylation of R-SMADs. It also associates with SMURFs to form the SMAD7-SMURF complex after TGF-Beta stimulation and ubiquitinates the receptors on the cell surface or endosomal membranes; these are then targeted for degradation in proteasomes and lysosomes (Ref.4). Another adaptor protein, STRAP1, also binds to both Type-I receptors and SMAD7, and enhances the inhibitory activity of SMAD7 (Ref.5).
Solun sisällä esiintyvät mikrotubulukset toimivat
raiteina solun sisäisille SMAD-proteiinien liikenteelle.
FILAMIINI, joka on aktiiniverkostoa ristiin rastiin tekevä
ja rakenteellinen proteiini, liittyy myös SMAD-proteiiniin ja
säätelee positiivisesti SMAD-signaalien johtamista.
SMAD-signalointia voi säätää Ras-ERK-MAPK- signaalitie
vasteena RTK-aktivaatiolle. SMAD-signalointia voi säätää myös
SMAD2:n toiminnallinen interaktio kalsiumkalmoduliinin kanssa.
Lisäksi inhibitoristen SMADien ( SMAD6 ja SMAD7)
ilmentymistä vahvistaa moninaiset signaalit kuten EGF
(epidermaalinen kasvutekijä), AP -1 (aktivaattoriproteiini 1)
ja IFN-gamma( interferoni gamma) ja nämä muodostavat tärkeän
mekanismin, jonka kautta nämä tiet voivat säätää
negatiivisesti SMAD-aktivaatiota.
-
SMAD signaling can be regulated by the Ras-ERK-MAPK pathway in response to receptor tyrosine kinase activation and also by the functional interaction of SMAD2 with Ca2+-Calmodulin (Ref.8). In addition, the expression of SMAD6 and SMAD7 is enhanced by multiple signals including EGF (Epidermal Growth Factor), stimulation of AP-1 (Activator Protein-1) by phorbol ester 12-O Tetradecanoylphorbol-13 Acetate, and IFN-Gamma (Interferon-Gamma), which provide an important mechanism whereby these pathways negatively regulate SMAD activation (Ref.9).
SMAD-proteiineja ilmentyy yleisesti koko kehityksen ajan ja kaikissa aikuiskudoksissa; monia niistä (SMAD2, SMAD4/DPC4, SMAD5, SMAD6 ja SMAD8) tuottuu alternatiivisesti pleissautuneista mRNA-molekyyleistä. Tärkeitä TGF-beetan transkriptionaalisissa antiproliferatiivisissa vasteissa ovat SMAD2 ja SMAD4/DPC4. Ja koska ne ovat ihmissyövissä inaktivoituneita, päätellään niiden olevan tuumorisuppressoreita. SMAD3-deleetio johtaa hitaaseen follikulaariseen kasvuun, lisääntyneeseen atresiaan ja infertiliteettiin joko hormonipitoisuuksiin vaikuttamalla ovariaalisten seletiivisten reseptorien muuntumisen takia ja /tai ovariaalisten solujen elossapysymistä säätelevien geenien muuntumisesta.
-
SMADs are ubiquitously expressed throughout development and in all adult tissues and many of them (SMAD2, SMAD4/DPC4, SMAD5,SMAD6 and SMAD8) are produced from alternatively spliced mRNAs (Ref.4). SMAD2 and SMAD4/DPC4 are important for transcriptional and antiproliferative responses to TGF-Beta, and their inactivation in human cancers indicates that they are tumor suppressors (Ref. 10, 11). Deletion of SMAD3 results in slow follicular growth, increased atresia, and infertility by either affecting selected hormone levels, by altering the expression of selected receptors in the ovary and/or by altering genes that regulate cell survival in the ovary.
Kartta
kuva inhibitorisesta SMAD7 proteiinista ja sen interaktioista,
joilla se blokeeraa TGF-beta signalointia
The role of Smad7
https://openi.nlm.nih.gov/detailedresult.php?img=PMC2538763_6604562f5&req=4
The role of Smad7
https://openi.nlm.nih.gov/detailedresult.php?img=PMC2538763_6604562f5&req=4
Inga kommentarer:
Skicka en kommentar