Repetition about interferons IFN and influenza virus
April 27, 2009
I. Lähinnä interferoneista vanhan kertausta
https://ars.els-cdn.com/content/image/1-s2.0-S1050464805000690-gr1.jpg
2. Miten A-influenssa vikuuttaa luonnollista immuunipuolustusta ja hävittää interferonivasteen.
LÄHDE: Adolfo Carcia-Sastre. Pathogenesis Antiviral Response in Pandemic Influenza Viruses Mount Sinai School of Medicine, New York, US
I. SOLUN INTERFERONIT ANTIVIRUSVASTEENA JA VIRUSTEN ANTI-INTERFERONIVASTE
Tämä newyorkilainen teksti pureutuu isäntäsolun antivirusvasteen selvittelyyn. Ihmisen solussa on paljon , yli 100 antivirusgeeniä. Mutta on kyse vain siitä, ehtiikö solu aktivoida ne antivirusgeeninsä vai ei ja saako solu kehkeytettyä järjestelmän propagoitumisen interferoneilla vai ei.
II. Kirjoituksen lopussa kirjoittaja käsittelee influenssavirusten suhtautumista tähän viruksia taltuttavaan IFN-järjestelmään. ”Influenssaviruksen aiheuttamat antivirusvasteet kehossa ja asian patogeneesi”.
Myös tulevaisuuden näkymiä antivirusstrategiasta.
I IFN-järjestelmä
Virusten ja ihmiskehon välisistä monimutkaisista vuorovaikutuksista riippuu virusinfektion lopullinen tulos. Virusinfektiot varsinkin ovat aivan erityisen päteviä kehon signaaliohjelmien esiinliipaisijoita infektoituneessa solussa. Interferoni on infektoituneen solun hätähuuto terveille soluille ympäristössä. Virusvaikutuksesta aiheutuu isäntäsolun (tässä: ihmissolun) geenien asennoituminen asiaan, moduloituminen, virittyminen ja geenituotteitten määrissä tapahtuu havaittavia muutoksia. Toiset näistä muutoksista suosivat viruksen omaa replikaatioprosessia (lisääntymistä) ja toiset muutokset heijastavat isäntäsolun vastetta virusta kohtaan.
Tässä tulee esiin VIRUSTEN pitkän evoluution ja kokemuksen tuloksena kyhätty viruksen todellista IHMISTUNTEMUSTA OSOITTAVA sopeutunut GEENIPÄTEVYYS. Ne ovat jo edeltä (evoluutionsa aikana) valmistautuneet kohtaamaan isäntäsolun luontaisen, normaalin, alkupuolustuksen. Tästä viruksen ja isäntäsolun keskinäisestä suhteesta, interaktiosta, syntyykin taudin asteittainen eteneminen – ”taudin tango” ja vaikutukset adaptatiiviseen immuunivasteeseen. Virus taistelee ikään kuin vertaisensa ”järkiapparaatin” kanssa mitä pätevimmin, mitä loogisimmin ja kauaskantoisin strategioin, jos ei muuten, niin koettamalla ”ehtiä ennen”. Tekstiin liittyvässä kuvassa virus koettaa ehtiä saamaan RNA-apparaattinsa nopeasti sytoplasman havaitsijajärjestelmien huomaamatta tumaan, ehkä DNA- muodossa ihmis DNA:n sekaan. Se käyttää aikatekijää hyödyksi, sillä viruksen vastainen järjestelmä vaatii ihmisen solun DNA:n asennoitumisen tuottamaan IFN, interferoneja. Mutta jos influenssavirus jo on muutamilta geeneiltään ja eväsproteiineiltaan tumassa se estää monella tavalla interferonijärjestelmän hienosäätöistä moniaskelista järjestelmää pääsemästä täyteen vauhtiin
Viruksen puolelta:
VIRUSPROTEIINIEN säätelyfunktiot ovat hyvin merkitseviä influenssaviruksen patogeenisyydessä ja ne näyttävät osittain olevan syynä influenssaviruksen kykyyn infektoida useitakin eläinlajeja, mistä sitten seuraa ihmiskunnalle asteittain kehkeytyvä pandemisen influenssan (uuden viruksen) vaara. Patogeenien kehittyminen rinta rinnan niiden isäntien kera on johtanut isäntäkehojen immuunijärjestelmien muovautumiseen, ehkä terveelliseen virittymiseenkin toisaalta.
Ihmisen tai muun isäntäkehon puolelta:
ISÄNNÄN PUOLELLA pääkomponenttina tässä järjestelmässä on LUONNOLLINEN IMMUUNIVASTE, johon kuuluvat kaikki isäntäkehon eri estemuodostumat, barrierit, kuten iho, limakalvot, limakalvojen eri eritteet ja monet patogeenispesifiset vasteet. Tämä luonnollinen immuunivaste ei käsitä vain isännän ensimmäistä infektioitten estettä, vaan varustaa kehoa myös signaaleilla, joista seuraa SOLUVÄLITTEISTEN ja HUMORAALISTEN IMMUUNIVASTEIDEN kehittyminen.
IFN, interferoni
Interferoni-molekyylien kehkeytyminen ihmissoulssa on merkkinä virusinfektiosta
(1) Interferonit IFN alfa ja IFN beeta ovat kriittinen elementti tässä luonnollisessa immuunijärjestelmässä. Nämä ovat erittäin tärkeitä kehon puolustautuessa varsinkin viruksilta. Kun erittyy näitä interferoneja, ne vaikuttavat solujen antivirusvasteen esiin kehkeytymisen lisääntymän säätämällä antivirusvasteen geenijärjestelmän käyntiin
(2) Interferoni beeta-geeni säätyy päälle. Lopulta voi transkription kautta yli 100 antiviraalia geeniä käynnistyä ja propagoida tehokkaan vasteen ja johtaa kehon tervehtymiseen ja viruksen voittamiseen.
VIRUS HANKKII evoluutiopakosta itselleen joskus vuosittainkin UUSIA GEENEJÄ selvitäkseen ihmisen paremmasta virusvasteesta.
On ymmärrettävää, että jos virus haluaa elää maapallolla, sen on päinvastoin kyettävä sammuttamaan tämä ihmisen tai eläimen antivirusjärjestelmä kokonaan tai osittain ainakin puolustukseen osallistuvista soluista. Huolimatta kehon ja solujen mitä hienosääteisimmästä immuunijärjestelmästä onnistuu virusten kuitenkin infektoida soluja ja aiheuttaa kehon sairauksia, joskus jopa kuolemaa. Tämä virusten saama menestys selittyy ainakin osittain niitten tehokkaasta evoluution aikana tapahtuneesta omien geenien hankkimisesta, joilla ne ovat oppineet VASTUSTAMAAN - tai VÄLTTÄMÄÄN, KIERTÄMÄÄN ja PETTÄMÄÄN - ihmisen IMMUUNIPUOLUSTUSTA. Jopa menossa olevan pandemian aikana virus voi muokata geenistöään tai mutatoitua virulentimmaksi.
VIRUS SÄÄTÄÄ PÄÄLLE IFN-järjestelmän VASTA-VAIKUTTAJIA
Virus säätää vauhtiin IFN-antagonisteja:
Erityisen kiinnostavia ovat viruksen koodaamat interferonien (IFN) vastavaikuttajat, IFNalfa/beeta-antagonistit, koska niitä näyttää sijaitsevan useimmissa animaalisissa viruksissa.
(Artikkelin kirjoittaja selittää lopussa yksityiskohtaisesti, miten influenssavirus pystyy välttämään eräällä ihmisen virusvastetta vastustavalla geenillään (NS1) ihmisen luontaisen immuunipuolustuksen ja tutkija kohdistaa tutkimuksensa erityisesti interferonivasteeseen soveltaen tätä tietoa mahdolliseen pandemiaan).
MIKÄ ON INTERFERONIEN NORMAALITOIMINTA VIRUSINFEKTIOSSA?
IFNα/β (interferonien alfa ja beeta) (1) vaikuttama antivirusvaste.
Kun eläinkehon tai ihmiskehon solu havaitsee virusinfektion, se alkaa aika välittömästi syntetisoida ja erittää interferoneja IFN alfa / beeta. IFNα/β-geeneihin kuuluu IFNβ-geeni ja monta sen lähisukuista IFNα-geeniä. Interferoni itse on rakenteeltaan glykoproteiini (gp). Säätely ei ole aivan suoraviivaista.
Yleensä tähän interferonien herättämistoimintaan osallistuu monta proteiinia (kaskadi). Sellainen virusmuoto, jossa on kaksoisrihma (dsRNA) on se mikä antaa hälytyksen tämän kaskadin kehkeytymiseeen.
Hälytyksen aihe tai sanoma : VIERAS ”dsRNA” TULLUT SOLUUN!!! (Kts. kuvaa viruksen vRNP paketista, jonka virus koettaa muiluttaa tumaan, niin että sytolasmassa ei havaittaisi sen RNA- muotoa:
Seuraavia tekijöitä alkaa yleensä normaalisti mistä tahansa virushälytyksestä herätä:
SENSORIT ovat (3) RIG-1 ja (4) MDA-5 sensoriproteiinit, helikaasit, ja ne tunnistavat yleensä virus dsRNA:ta sytoplasmasta.
(3) Helikaasi RIG-1 = DExD/H (Asp-Glu-Ala-Asp/His) box polypeptide RIG-I.
(4) Helikaasi MDA-5 = ”An interferon-inducible putative RNA helicase”. ”MDA-5 on dsRNA:sta riippuvainen ATPaasi, jossa on sekä kaspaasia rekrytoiva domeeni että RNA-helikaasimotiivi. ( a double-stranded RNA-dependent ATPase that contains both a caspase recruitment domain and RNA helicase motifs).
(5,6 ) ADAPTORIT: ovat IPS-1 ja MAVS, eräitä solutekijöitä, jotka alkavat aktivoitua edellisten jälkeen.
(5) IPS-1 = adaptori, joka vaikuttaa kahden edellämainitun tekijän välittämässä antiviraalissa vasteessa; (an adaptor involved in RIG-I- and Mda5-mediated antiviral immune responses). (IPS-1= ”interferon inducers, a molecule interferon-beta promoter stimulator 1), interferonin indusoija, IFN-beetapromoottorin stimulaattori-1. Lisänimikin on: (IPS-1 = ”Putative NF-kappa-B activating protein”) Nat Immunol. 2005 Oct;6(10):981-8.
(6) MAVs = antiviraali-signaloiva –valkuaisaine, joka aktivoi NF-kB:n ja IRF3:n. (”a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF3. Mitokondriaalinen antiviraalinen signaloiva proteiini, joka aktivoi NF-kB: ja IRF3 tekijät.
MAV proteiineilla voi olla ihmisen immuunivasteessa suuri merkitys.
(7) AKTIVOIVAT FOSFORYLOIJAT: IRF-kinaasit tulevat kuvaan. Nämä kinaasit ovat fosforyloivia
(7.1.) TBK1 = TANK-binding kinase 1
(7.2.) IKKε jota tarvitaan NF-κB aktivaatioon T-solu reseptoreilla (TKR)
TUMATEKIJÄT
(8) NF-κB =Nuclear Factor kB, tumatekijä kB, transkriptiotekijä
( Jos IKB-kinaasit fosforyloivat IKB proteiinien seriinitähteitä, ne merkkautuvat proteiinisilppuritiehen , ubikitinylaatioon ja silloin salliutuu NF-kB-kompleksin aktivoituminen ja siirtyminen tumaan. Phosphorylation of serine residues on the IKB proteins by IKB kinases marks them for destruction via the ubiquitination pathway, thereby allowing activation and nuclear translocation of the NF-kB complex).
MUITA TRANSKRIPTIOTEKIJÖITÄ
(9) AP-1. Tumaan siirtyy sytoplasmasta DNA:lle AP-1 = transkriptiotekijä, ja tekee c-jun/c-fos dimeeri kompleksin DNA:n kanss
KOHDEGEENI
(2) IFN-beeta-geeni indusoituu ja sen transkriptio alkaa: Mitä syntyy?
MUODOSTUU TÄRKEÄ mRNA, JOKA VOI ANTAA OHJELMAN INTERFERONIEN TUOTOSTA
(10) Pre-mRNA isäntäsolun omasta DNA:sta sairastuneen isäntäsolun tumassa.
(11) Pre-mRNA modifioidaan (Asetetaan suuntajana CAP ja polyadenyloidaan-AAAAA-) ja lähetetään sytoplasmaan, jossa syntyy IFN-beeta.
Siis—-
KUN JOTAIN VIERASTA VIRAALIA AINESTA HAVAITAAN ( dsRNA muoto on varoitusmerkki). on monella solulla solunsisäisiä virustuotteitten havaintolaitteita, ”sensoreita” ja kun sellainen aktivoituu, se aloittaa signalointisarjan, kaskadin, joka johtaa solun omassa tumassa sijaitsevan transkription kautta tapahtuvaan IFNβ-geenin aktivoitumiseen (2) ja interferonin tuottoon.
HELIKAASIT ovat viime vuosikymmeninä löytyneitä molekyylejä. Ne ovat niitä sensoreita. Näitten sensorien laatua ei aiemmin tunnettu, mutta nyt tiedetään, että on kyse kahdesta RNA-helikaasista, RIG-1 ja MDA-5, jotka tuntevat viruksen dsRNA- muodon ja ne helikaasit syntyvät sytoplasmassa virusinfektion aikana.
Kun nämä helikaasit RIG-1 (3) ja MDA-5 (4) sitoutuvat vieraisiin dsRNA-rihmoihin, alkaa niitten helikaasiaktiviteetti toimia ja seuraa struktuurin muutos, joka rekrytoi paikalle muita solutekijöitä kuten (vastikään keksityt) IPS-1/ MAVS-proteiinit (5 ja 6). Sitten aktivoituvat solun erilaiset kinaasit (7) (Kinaasien tehtävä on fosforyloida proteiineja). Näitten kinaasien joukossa on IRF3-kinaasit TBK1 ja IKKε, jotka aktivoituvat (7.1. ja 7.2.) . Aktivoituneet IRF3-kinaasit yhdessä tumatekijän NF-κB (8) and (9) transkriptiotekijän AP-1- kanssa kertyvät tumaan. Ne sitoutuvat ihmisen tai eläimen DNA:ssa sijaitsevaan (2) interferonigeeniin IFNβ, sen promoottoriin ja stimuloivat siitä isäntäsolun alustavan mRNA:n esiintuloa (transkriptiolla interferonigeenistä). Näin saadaan lopulta kypsää interferoni mRNA:ta.
Minkä tahansa ihmisolun sytoplasmaan tullut viraali dsRNA-muoto on sellainen virusmolekyyli, joka liipaisee tämän ylläkuvatun kaskadin esiin. Lienee myös muita virustuotteita ja muita solusensoreita, jotka yhtä hyvin kykenevät osallistumaan IFNβ-geenin induktioon.
ON TOINENKIN TAPA, MITEN INTERFERONIERITYS VOI LÄHTEÄ KÄYNTIIN
Tyypin I IFN signalointi
Jos virusperäistä RNA- tai DNA-materiaalia on endosomissa, voi IFNα/β-induktio myös stimuloitua käyntiin seuraavaa tietä:
TL- Reseptorit TLR3, TLR7, TLR8 tai TLR9 osallistuvat signalointiin. Eri solulajit näyttävät käyttävän erilaisia mekanismeja virusten tuotteitten tunnistamisessa.
(12) TLR3, TLR7, TLR8 tai TLR9 (Toll-like receptor)
MITÄ SYNTYNEET INTERFERONIT (IFN) TEKEVÄT?
Kun interferoneja (IFNα/β) on sitten syntetisoitunut sairaassa solussa, niitä eritetään infektoituneesta solusta ja ne kertovat ympäristöön, että solulla on hätätilanne. Niitä asettuu niille spesifisiin reseptoreihin, (13) vastaanottajiin (IFNα/β receptori)
Kaikki alfa- ja beeta-interferonit kiinnittyvät samanlaiseen reseptoriin.
NE HERÄTTÄVÄT TERVEISSÄ SOLUISSA ANTIVIRUSVASTEEN.
Eritetyt interferonit sitoutuvat reseptoreihin lähisolujen pinnalla ja aiheuttavat niistä antiviraalisten proteiinien tuottoa, jolloin kohdesolut tulevat resistentimmiksi virusta vastaan.( mutta solun täytyy ehtiä kehittää sanomaa lähisoluille - Juuri tässä virukset koettavat voittaa aikaa ja estää tiedonannon jolloin solu toisensa jälkeen nääntyy yksin ja lopulta virus on voittanut koko immuunipuolustuksen)
Kun IFN on sitoutunut solupintareseptoriinin, terveen solun reseptorissa, solunliman puolella oleva osa aktivoituu ja aktivoi solun sisällä JAK-kinaasin (14), erään tyroosiinikinaasin.
(14) Janus activated kinase, tyrosiinikinaasien ryhmää).
Tämä taas katalysoi STAT-fosforylaation.(15)
(15) STAT on yleensä inaktiivina sytoplasmassa sijaitseva transkriptiotekijä.
STAT1 ja STAT2 tarkoittavat ”Signal Transducers and Activators of Transcription”.
STAT dimerisoituu, siirtyy solulimasta tumaan, sitoutuu tuman asiaankuuluvaan DNA-response elementtiin ja aktivoi asiaankuuluvan transkription ja silloin alkaa tapahtua.
JAK-STAT-aktivaatio on spesifinen.
Eri tyypin IFN aktivoi eri tyypin STAT-molekyylin ja täten eri kohtia DNA:sta tulee aktivoitua.
Muutkin signaalimolekyylit kuin IFN voivat saada STAT- fosforylaation aikaan.
Vain STAT ja CREB tunnetuista transkriptiotekijöistä ovat sellaisia, jotka aktivoituvat fosforylaatiosta.
(16) Ennenkuin STAT menee tuman puolelle, se on muodostanut molekyylikompleksin, jonka nimenä on ISGF3 = interferonillastimuloitu geenitekijä 3 ( complex of STAT1, STAT2 and IRF9). Se on ligandista riippuva transkriptionaalinen aktivaattori, joka vasteena interteronikäsittelylle koostuu kokoon solun sytoplasmassa, sitten translokoituu tuman puolelle ja sitoutuu consensus-DNA-kohtaan interferonilla stimuloituvaan elementtiin ISRE.
(Interferon-stimulated gene factor 3 (ISGF3) is the ligand-dependent transcriptional activator that, in response to interferon treatment, is assembled in the cell cytoplasm, is translocated to the nucleus, and binds the consensus DNA site, the interferon-stimulated response element)
(17) ISRE
Tumassa aktivoitunut transkriptiotekijä, kompleksi ISGF3, sitoutuu solun oman DNA:n promoottoriin, jossa on interferonilla stimuloituvia vaste –elementtejä (IRSE response element REs) ja näin alkaa mennä päälle, käynnistyä, monenlaisia antivirusgeenejä transkriptioteitse- normaalisti. Niitähän on yli 100 geeniä.
http://www.nature.com/nrmicro/journal/v7/n2/images/nrmicro2071-i3.jpg
(18) ANTIVIRUSGEENEJÄ
Näihin antivirusgeeneihin kuuluu MxA, PKR, OAS, ADAR, PML, p56 ja monta muuta, yli 100 geeniä. Nämä interferoni (IFN)-teitse käynnistetyt geenit estävät viruksen replikaatiota monella eri mekanismilla,
esim. sitoutumalla viruksen nukleokapsidiin
estämällä virus mRNA:n kautta tapahtuvaa translaatiotoimintaam (viruksen omien proteiinien tuottoa),
hajoittamalla RNA:ta,
muuntamalla RNA:ta
tai indusoimalla apoptoosia.
(MxA, MX1 geeni, joka koodaa IFN-indusoimaa p78 proteiinia (MxA).
www.uniklinik-freiburg.de/virologie/live/forschung/teams/hallerteam/MxProteins/mxa_domains.gif
PKR= dsRNA-dependent Protein Kinase, RNA:sta riippuva proteiinikinaasi
www.biochemsoctrans.org/bst/034/0007/bst0340007u01.gif
ja siitä riippumatonkin tie on :
www.cellscience.com/reviews6/Corbett3.jpg
ADAR= adenosine deaminase 1 (ADAR 1), adenosiinideaminaasi1
PML = promyelocytic. leukemia protein (PML)
OAS = oligoadenylate synthetase, oligoadenylaattisyntetaasi
p56 = Interferon-inducible protein p56, interferonilla indusoituva proteiini p56, translaation inhibiittori. Estää viruksen infektoiman solun translaatiotapahtumat.
”P56 binds to the translation initiation factor eIF3 through the subunit p48/Int-6. Together with the protein kinase PKR and the 2-5(A) synthetase, p56 downregulates translation in response to double-stranded RNA, e.g. in virus-infected cells. The 2-5(A) oligonucleotides activate RNase L. Unlike PKR and 2-5(A) synthetase, p56 does not require double-stranded RNA for activity and is therefore already active before cells are infected”) .
INTERFERONIEN (IFN) MUITA TEHTÄVIÄ
Lisäksi erittyneet interferonit (IFNα/β) edistävät vahvan SOLUVÄLITTEISEN ja HUMORAALISEN IMMUNITEETIN kehkeytymistä. Yleisesti ottaen interferonivasteella on monimutkainen säätyminen, mihin kuuluu positiivisia ja negatiivisia feed back -(takaisinsyöttö) ilmiöitä, joista osa on vielä tuntemattomiakin.
II
A Influenssavirus ja sen strategia ihmisen interferonijärjestelmää vastaan
Nyt artikkelin kirjottaja tarkkaa A-influenssavirusta, miten se suhtautuu interferonijärjestelmään, joka helposti voisi tuhota sen, jos ihmiskeho vain ehtisi ja voisi aktivoida kaikki 100 geeniä ja interferonituoton..
Influenssavirus ei lähde ”soitellen sotaan”. Sillä on ”eväänä” aika monta NS1 proteiinia virioninsa sisällä ja myös katteena viruspartikkelille , ”aivojuntalleen (vRNP)", mikä estää tämän ssRNA materiaalin havaitsemista solun puolustuksen sensoreissa.
NS1 JA GEENISEGMENTTI 8.
Mikä on tämä NS1, virusgenomin segmentti 8:n koodaama virusproteiini, nonstrukturaalinen proteiini?
Jo varhain on havaittu, että influenssavirukset ovat niukkoja interferonien indusoijia, vaikka interferonit kuvattiinkin ”antivirustekijöinä” ensimmäisen kerran soluista, joita oli käsitelty heikennetyillä, kuumakäsittelyllä inaktivoiduilla A-influenssaviruksilla.
Influenssavirukset, kuten moni muukin virus pystyy koodaamaan sellaisia mekanismeja, joilla voi VÄLTTÄÄ interferonivasteen tai VASTUSTAA sitä.
Mitä A-influenssa-virukseen tulee, sillä on oikein asialle antautunut antagonistifunktiota tekevä GEENI, jonka tuotteena on yllämainittu NS1-proteiini, nonstrukturelli proteiini1. Influenssaviruspartikkelissa on ainakin 8 eri palasta, jaksoa, segmenttiä RNA-materiaalia. Kahdeksas segmentti on lyhin ja siinä on se geeni, josta virus saa valmistettua näitä NS-proteiinia. Niitä on NS1 ja NS2.
NS1 proteiini on se nonstrukturelleista proteiineista, jota influenssaviruksella infektoituneessa ihmissolussa on eniten.
JOS NS1 GEENI PUUTTUISI VIRUKSELTA, MITÄ SITTEN?
Kun tehtiin tutkimuksia poistogeenisella tekniikalla ja kehitettiin influenssavirus, jossa ei ollut tätä NS1-geeniä tai tehtiin NS1-mutantti virus, havaittiin seuraavaa: Jos NS1 puuttui, viruksen replikoituminen oli puutteellista useimmissa soluissa ja isännissä, PAITSI jos isäntäsolusta puuttui toimiva interferonijärjestelmä. Mutantti NS1 virus oli heikentynyt, mutta kykeni suorittamaan replikaatiota.
ENTÄ JOS ISÄNTÄSOLUSTA PUUTTUU STAT 1 ja 2 (INAKTIIVI TRANSKRIPTIOTEKIJÄ)?
Sellaisessa koe-eläimessä, jolta puuttui interferonivasteen eräs avainmolekyyli STAT1 (STAT1 poistogeeninen koe-eläin), mutantti NS1 virus aiheutti taudin. Näistä tuloksista voitiin päätellä, että virus tarvitsee välttämättä NS1 geeniään ja geenituotetta ( NS1 proteiinia) voittaakseen isäntäsolun interferonivasteen influenssa A-virusinfektion aikana.
Influenssa A-viruksen NS1 geenin (Segm.8) IFNalfa ja beeta- antagonisoiva vaikutus perustuu NS1 proteiinin kykyyn estää IFNbeeta-synteesi. Tästä seuraakin A-influenssa viruksen maine, että se on sellainen ”huonosti beeta-interferoneja indusoiva virus”.
JOS NS1 PUUTTUU?
Kun ja jos NS1 puuttuu, influenssa A virus muuttuu vahvasti IFNα/β-interferoneja indusoivaksi kuten tavallinen virus yleensä – ja tällainen taas ehkäisee NS1-mutanttien virusten replikoitumista.
NS1 SITOUTUU ITSE ds-RNA MUOTOIHIN! ( Kuin olisi viruksen puolesta sensorina)
Koska NS1 pystyy sitoutumaan viruksen oman replikaation välituotteeseen, dsRNA-muotoon, on todennäköistä, että se sakkaa viruksen tuottamia dsRNA-rihmoja virusinfektion aikana, mikä taas estää tämän virusta haittaavan ja infektiota paljastavan muodon joutumista isäntäsolusensorien havaintokenttään.
(Tappaja-T-solut heräisivät myös).
Tämä toimintamalli sopisi kuvaamaan NS1 expression kyvykkyyttä estää transkription aktivoituminen siltä osin, mikä koskee interferonien IFNα/β synteesin indusoitumista IRF3 mukaanluettuna.
TUMA EI ANNA VIRUSHÄLYTYSTÄ
Lisäksi interferoni beetan (IFNβ) tuotto estyy virukselle optimaalisella tavalla silloin kun NS1-proteiini sitoutuu dsRNA-molekyyliin.
B-INFLUENSSA JA A-INFLUENSSA EROAVAT TOISISTAAN
B-influenssan suhteen on havaittu samankaltaisia löytöjä NS1 proteiinista. Kuitenkin NS1-proteiinin interaktiot soluproteiinien kanssa todennäköisesti vaikuttavat osaltaan myös sen interferoneja antagonisoivaan funktioon.
NS1 ESTÄÄ NORMAALIN mRNA:n prosessoimista vain A-INFLUENSSASSA.
www.nature.com/emboj/journal/v18/n8/images/7591652f7.jpg
Influenssa A-viruksen NS1 (mutta ei influenssa B-viruksen NS1) estää niitä solutekijöitä, jotka osallistuvat solun tavallisen mRNA:n prosessoimiseen. Tämäkin funktio saattanee omata osuutta A-viruksien interferonien tuoton estymisessä.
www.expasy.ch/viralzone/all_by_species/80.html
INTERFERONIEN ESTOVAIKUTUSTAKIN
Lopuksi mainittakoon että NS1 on osoittanut myös interferoneja inhiboivaa vaikutusta niitten jo ollessa syntetisoituneina ( interferonien postsynteesivaiheessa).
TRANSLAATIONESTÄJÄN AKTIVAATIO ESTETÄÄN.
Sekä A- että B-influenssavirukset estävät translaatioinhibiittorin ja interferoneja indusoivan proteiinin PKR aktivoitumisen. (Viruksen oma translaatio tapahtuu sytoplasmassa viruksen mRNA(+) :n avulla käyttäen solun koneistoa ja sillä tuotetaan virusproteiineja).
ANTIVIRAALISTEN VASTEIDEN KIIHDYTTÄJÄN ESTÄMINEN.
B-viruksen NS1 estää ISG15 aktiviteettia. Tämä taas on interferonien indusoima proteiini, jonka tulisi kiihdyttää interferonivälitteisten antiviraalisten vasteitten esiintuloa.ISG15 omaa luonnollisessa immuniteetissa osuutta ja se on ubikitiinin kaltainen proteiini.
(ISG15: ISG15 has been something of a mystery to biologists, and nobody has known … However, scientists have reasoned that ISG15 is involved in the innate immune . ISG15 (IFN-stimulated gene, 15kDa) proteins is a UBl (ubiquitin-like protein)
NS1 geenin osuus ( Virusgeeni 8) virulenssissa
Ihmisinfluenssa A virus pystyy infektoimaan suoraan monia eri eläinlajeja, kuten eri lintuja (esim.vesilintuja, kananpoikia, kalkkunoita), hevosia, porsaita ja ihmisiä, mutta se voi myös ylittää lajirajoja, kuten lintuinfluenssavirus ja sikainfluenssa.
Avian influensavirus A voi infektoida nisäkäseläimiä ja myös ihmistä.
http://avianflu.unair.ac.id/images/ekologi3.jpg
Tästä ominaisuudesta on seurannut historian aikana kriittisiä ihmiskunnan pandemioita, joille on ominaista jokin uusi antigeeninen piirre, uusia virusdeterminantteja, uuden viruksen lähtö kiertämään maapalloa. Nämä determinantit taas ovat peräisin sellaisista lintuviruskannoista, joille ei ole olemassa edeltäkäsin mitään spesifistä, osuvaa immuniteettia useimmissa ihmisväestöissä. Tällöin uudesta viruksesta olisi tuloksena korkempi sairastuvuus ja mortaliteetti, kuin tutumman vanhan ihmisinfluenssaviruksen palatessa. Tosin on vaikea ennustella mitään linnuissa kiertelevien influenssaviruskantojen pandemisista potentiaaleista. Eihän tunneta täydellisesti niitä tekijöitä, jotka osallistuvat lintuinfluenssakantojen tai lintuantigeenideterminantteja sisältävien uudelleen järjestyneitten virusten kykyyn infektoida ja lisääntyä (propagoida, riehahtaa) ihmisissä. Tämän tiedonpuutteen takia ei tarkkoja pandemisen potentiaalin arvioita ole annettavissa. (Influenssa A viruksen siirtymä välillä sikoihin on kuin virus olisi asettunut tehokaaseen vispilään muokkaamaan kapasiteettejaan).
HA GEENI ja HEMAGGLUTINIINI
Vaikka näyttää selvältä, että hemagglutiniiniproteiinin (HA) reseptorispesifisyys on tärkeä seikka lintuviruskantojen sopeutumisessa ihmisiin, on kuitenkin muita, vähemmän selviteltyjä tekijöitä, jotka osallistuvat tähän lajirajan ylittävään ihmisiin sopeutumiseen, adaptaatioon.
NS1 GEENI VASTAVAIKUTTAA INTERFERONIJÄRJESTELMÄÄN
Mitä tulee NS1-geeneihin, niin eri viruskannat eri eläinlajeista näyttävät valitsevan todennäköisimmin sellaisen NS1-geeniadaptaation, jonka tuloksena saadaan aikaan juuri sen lajin interferonijärjestelmän (IFNα/β system) lajispesifinen antagonismi, vastavaikutus.
H1N1/NS1/1918/WSN ihmisinfluenssa, hiiren influenssa A/H1N1.
Kun vertailtiin vuoden 1918 pandemian (Espanjan taudin A-influenssaviruksen H1N1) NS1-geeniä hiireen sopeutuneen A/H1N1-influenssaviruskannan NS1-geenin kanssa, havaittiin tapahtuneen juuri kuten yllä mainittiin.
(Kyse oli A-influenssavirus kannasta WSN). Kussakin lajissa NS1 geeni sopeutuu sille lajille tyypillisesti vastustamaan juuri sen lajin interferonijärjestelmää. Kun sijoitettiin WSN virukseen (WSN = Wild type influenza virus) sen NS1-geenin kohdalle vuoden 1918 viruksen NS1-geeni, havaittiin, että virus oli vaimentunut hiiressä, mutta ihmissoluissa pystyi aiheuttamaan tehokkaan interferonituoton estymisen. Asian selittänee NS1 geenin spesifiset interaktiot isäntätekijöihin, joissa on lajista riippuen erilaisia sekvenssejä. Joten hiireen sopeutunut NS1 oli vahvemmassa interaktiossa hiirien eri tekijöitten kanssa kuin ihmisen eri faktoreitten kanssa ja päinvastoin.
Jos tämä on eri influenssaviruskantojen NS1-geenin yleinen ominaisuus, olisi lintuviruskannan NS1-geenin sopeuduttava siten, että se tehokkaasti antagonisoi ihmisen interferonijärjestelmää (The human IFNα/β system).
Vaihtoehtoisesti lintuviruksen olisi välttämättä hankittava joltain ihmisinfluenssakannalta päin NS1 geenin uudelleen järjestäytynyt (REASSESMENT) muoto, jotta se voisi tehokkaasti infektoida ja propagoida ihmiskunnan puolella.
NS1 GEENI ON MERKITSEVÄ
Koska mutaatiot, jotka koskettavat NS1-geenin funktiota, vaikuttavat vahvasti viruksen patogeenisuuteen, korkeasti patogeenisella influenssaviruskannalla (HPAIV) saattaa olla sellainen NS1-geeni, joka on ominaisuuksiltaan erityisen vahva interferonien (IFNα/β) vastavaikuttaja, antagonisti.
NS1-GEENI VAIMENTAA MUITAKIN TRANSKRIPTIOTEKIJÖITÄ VAIKUTTAEN SYTOKIINIEN ERITTYMISEEN. Edelleen: NS1-geenin kyky vaimentaa erilaisten transkriptiotekijöitten aktivoitumista virusinfektion kuluessa antaa muitakin vaikutuksia kuin vain edellä mainitut vaikutukset interferonien (IFNα/β) synteesiin
ERÄÄN HAITALLISEN (PROINFLAMMATORISEN) SYTOKIININ ERITYS VOI LISÄÄNTYÄ LIIKAA (TNF-alfa)
Esimerkiksi NS1 näyttää säätelevän dendriittisolujen funktion aktivoitumisessa mukana olevan monen sytokiinin ja molekyylin esiintymistä. Siltä osin E-Aasian siipikarjassa ja vesilinnuissa riehuvien vahvapatogeenisten (HPAI) lintuinfluenssavirusten NS1 voisi olla syynä näitten virusten aiheuttamaan ihmisten makrofagien kohonneeseen proinflammatoristen sytokiinien ( esim. TNF-alfa) pitoisuuteen. Proinflammatoristen sytokiinien korkeat määrät ovat todennäköinen syy näitten virusten ihmiskunnassa aiheuttamaan epätavalliseen letaalisuuteen. Onneksi näitten virusten aiheuttamat infektiot näyttävät olevan harvinaisia eikä H5N1 lintuinfluenssavirus ole osoittanut kykyjä propagoida tehokkaasti ihmisestä toiseen.
MUITAKIN VIRUSGENOMIN OSIA SYYLLISENÄ KEHOPUOLUSTUKSEN LUHISTUMISEEN
www.nature.com/scitable/nated/content/26874/10.1038_nrmicro1208-f1_mid_1.jpg
Viruksen PA, Geeni 3. Endonukleaasi
Vaikka NS1 olisi deletiogeeninään ja indusoisi runsaasti interferoneja alfa ja beeta, niin lisäksi UV-sädetys saisi vielä enemmän interferonieritystä esiin, joten täytyy olla lisäksi jokin muu geeni kuin NS1 yksin, mikä pystyy lopettamaan IFNalfa/beeta tuoton. Mahdollisesti viruspolymeraasi endonukleaasina, joka nappaa CAP-palasen (”cap-snatching”) solun omalta mRNA:lta, lie vastuussa muusta anti-IFN-alfa/beeta vaikutuksesta. Mutta tästä tarvitaan lisätutkimuksia. Joka tapauksessa useissa virusperheissä on tavallista monien IFNalfa/beeta antagonisoivien ko-operatiivisten virusgeenien olemassaolo.
Viruksen PB1 (ja PB1-F2), Geeni 2 polymeraasi, elongaasi (ja mitokondrian apoptoosiherkistäjä)
www.mcb.uct.ac.za/tutorial/myxo.gif
Influenssa-A-virus koodaa toista nonstrukturellia polypeptidiä viruksesta infektoituneissa soluissa ja se on PB1-F2 proteiini. Se koodautuu vaihtoehtoisella ORF-luennalla PB1-F2 segmentistä käsin, mikä myös ohjelmoi PB1-proteiinin synteesiä. PB1 taas on viruspolymeraasin kriittinen komponentti.
PB1-F2 löydetään infektoituneen solun mitokondriasta, missä se vaikuttaa mitokondrian läpäisevyyteen ja sen kalvoaukkokomplekseihin (ANT3, VDCA1). Näillä tekijöillä on tärkeä osuus apoptoosin kontrollissa. Tätä kautta PB1-F2 herkistää isäntäsolun apoptoosille. Jos influenssaviruksen PB1-F2 tekijä olisi tehoton, säästyisi isäntäsolu (ihmisen monosyytti) apoptoosilta. Onkin useita ihmisinfluenssakantoja, joista puuttuu tämä PB1-F2, joten sen olemassaolo saattanee vaikuttaa patogeenisyyteen ja sillä voi olla pandemian vaikeusasteessa merkitystä.
TULEVAISUUS: NS1 antiviruslääkkeitten ja rokotteen maalitauluna
INTERFERENSSI (RNAi, siRNA). Tietämyksellä NS1-geenin ja proteiinin funktioista voi olla sovellutusta lähitulevaisuudessa etsittäessä uusia anti-influenssaviruslääkkeitä. Jos löydettäisiin pieniä molekyylejä (siRNA) , jotka voisivat vaikuttaa interferenssiä NS1-tekijän kykyihin sitoa dsRNA:ta tai sen kykyyn estää IFNalfa/beetatuottoa, voitaisiin kohottaa isäntäkehon luonnollista immuniteettia influenssavirusta kohtaan ja keho selviäisi nopeammin viruksesta.
GEENIVAIMENNETTU ROKOTE. Lisäksi sellaiset rekombinanttivirusrokotteet, joissa olisi toimimaton NS1-geeni, saattaisivat edustaa tehokasta elävää heikennettyä rokotetta influenssavirusta vastaan. Tällaisia viruksia pystyttäisiin kasvattamaan sellaisissa elatusaineissa, missä ei ole IFNalfa/beetaa, mutta ne häviäisivät kehossa.
GEENIMODIFIOITU VIRUSROKOTE. Edelleen koska NS1 geenin estovaikutus kumoaa sekä luonnollista että adaptatiivista immuniteettia , olisi ilmeisesti mutantit NS1 virukset paljon immunogeenisempiä.
On kehitelty rekombinantteja influenssaviruksia, joissa on modifioidut NS1-geenit ja eri eläinkokeissa on todettu että ne ovat immunogeenisiä ja kumoutuvat vaikutukseltaan. Tällaisia modifioituja viruksia saatettaisiin käyttää tulevaisuudessa perustana eläville rokoteille epidemioita ja pandemista influenssaa vastaan.
This research was supported by the National Institutes of Health and Department of Defense. Dr García-Sastre is a professor in the department of microbiology at Mount Sinai School of Medicine in New York. His studies provided the first description and molecular analysis of a viral-encoded IFN antagonist among negative-strand RNA viruses.
Päivitystä27/04/2009 12:59
Suom. 2007-02-02 23:06
LÄHDE www.cdc.gov/ncidod/EID Emerging Infectious Diseases
Siirto Blogiin

onsdag 11 november 2009
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
onsdag 24 juni 2009
SLL kirjoittaa geeniluennasta
Suomen Lääkärilehdestä sitaatti:
Geeniluennan ymmärtäminen on avaintekijä tutkittaessa syövän syntyä
24.06.2009
Solujen stressinsietokyky on kohonnut monissa syöpäkasvaimissa. Syöpäsolut selviävät tällöin tilanteissa, joissa normaalit solut eivät pärjää: ne on ikään kuin ohjelmoitu jatkuvaan stressiin. Esimerkiksi säteily, raskasmetallit tai bakteeri- ja virustulehdukset ovat solustressiä, jotka voivat laukaista joko kuoleman mekanismin tai puolustusreaktion.
– Syöpäsolut tuottavat puolustusproteiineja, lämpösokkiproteiineja, suurentuneita määriä ilman ulkoisia ärsykkeitä. Solun oma puolustusmekanismi on erittäin riippuvainen siitä, kuinka hyvin solu voi, onko kyse nuoremmasta vai vanhemmasta solusta ja kuinka paljon vaurioita tapahtuu yhtä aikaa, kertoo professori Lea Sistonen Åbo Akademista.
Jotkut syöpäsolut eivät kuole solumyrkkyjen ja säteilytyksen jälkeen, vaan kestävät ne paremmin kuin normaalit solut. Myös tähän vaikuttaa stressiensietokyky.
Syöpäsolujen löydettävä oikea rakenne
Koska syöpäsolujen proteiinien tuotanto on lisääntynyt, proteiinien pitää löytää oikea kolmiulotteinen rakenne, eli laskostua oikein. Syöpäsolut tarvitsevat enemmän laskostumista auttavia proteiineja.
– Tällaiset molekyyliproteiinit toimivat ikään kuin “esiliinoina”. Ne sitoutuvat juuri syntetisoituneisiin polypeptideihin, jotka vapautuvat ribosomeista. Monesti juuri valmistuneiden proteiinien vettä hylkivät osat ovat auki. Tämä on kriittinen vaihe, sillä proteiinit saattavat mennä sikinsokin ja voivat muodostaa komplekseja väärien proteiinien kanssa. Suojelevat esiliinaproteiinit pitävät ne sellaisessa tilassa, etteivät ne pääse laskostumaan väärin, huomauttaa Sistonen.
Syöpäsolussa on ilmeisesti laskostukseen erikoistuneita suojeluproteiineja jo valmiiksi paljon, jotta ne selviävät syöpäsolun kiihtyvästä aineenvaihdunnasta.
Geeniluenta selvittää hiljentyvät ja aktivoituvat geenit
Lea Sistonen tutkii geeniluentaa, jossa selvitetään, mitkä geenit hiljennetään ja mitkä aktivoituvat. Ainoastaan murto-osa kaikissa soluissa olevista samoista geeneistä aktivoituu tietyissä vaiheissa.
– Haasteita aiheuttaa muun muassa se, että geeniluenta yleensä hiljennetään silloin, kun solut alkavat tuottaa suojelevia proteiineja. Vain tietyt lämpösokkitekijät aktivoituvat. Estäjiä ja stimulaattoreita etsitään nyt kovasti. Tutkimme parhaillaan sitä, miten geeniluenta tapahtuu ja kuinka sitä pitäisi joskus estää syövässä. Jos soluissa on yliaktiivinen stressivaste, se pitää hiljentää, kertoo Lea Sistonen.
Kolikon toinen puoli on se, että esimerkiksi Alzheimerin taudissa ja Parkinsonin taudissa aivoihin kertyy ei-toiminnallisia proteiineja, jotka pitäisi poistaa, mutta jotka eivät jostakin syystä häviä. Siksi nämä proteiinit sakkautuvat aivoihin.
– Toisissa soluissa siis pitäisi edesauttaa stressinsietokykyä, kun taas joissakin soluissa sitä pitäisi estää. Nyt tutkitaankin eri lääkeaineita, joista toivotaan apua siihen, että syöpäsoluihin voitaisiin viedä estäjiä ja aivoihin stimulaattoreita, painottaa Sistonen.
LÄHDE: UllaToikkanen
http://www.laakarilehti.fi/uutinen.html?opcode=show/news_id=7515/news_db=web_lehti2006/type=1
Geeniluennan ymmärtäminen on avaintekijä tutkittaessa syövän syntyä
24.06.2009
Solujen stressinsietokyky on kohonnut monissa syöpäkasvaimissa. Syöpäsolut selviävät tällöin tilanteissa, joissa normaalit solut eivät pärjää: ne on ikään kuin ohjelmoitu jatkuvaan stressiin. Esimerkiksi säteily, raskasmetallit tai bakteeri- ja virustulehdukset ovat solustressiä, jotka voivat laukaista joko kuoleman mekanismin tai puolustusreaktion.
– Syöpäsolut tuottavat puolustusproteiineja, lämpösokkiproteiineja, suurentuneita määriä ilman ulkoisia ärsykkeitä. Solun oma puolustusmekanismi on erittäin riippuvainen siitä, kuinka hyvin solu voi, onko kyse nuoremmasta vai vanhemmasta solusta ja kuinka paljon vaurioita tapahtuu yhtä aikaa, kertoo professori Lea Sistonen Åbo Akademista.
Jotkut syöpäsolut eivät kuole solumyrkkyjen ja säteilytyksen jälkeen, vaan kestävät ne paremmin kuin normaalit solut. Myös tähän vaikuttaa stressiensietokyky.
Syöpäsolujen löydettävä oikea rakenne
Koska syöpäsolujen proteiinien tuotanto on lisääntynyt, proteiinien pitää löytää oikea kolmiulotteinen rakenne, eli laskostua oikein. Syöpäsolut tarvitsevat enemmän laskostumista auttavia proteiineja.
– Tällaiset molekyyliproteiinit toimivat ikään kuin “esiliinoina”. Ne sitoutuvat juuri syntetisoituneisiin polypeptideihin, jotka vapautuvat ribosomeista. Monesti juuri valmistuneiden proteiinien vettä hylkivät osat ovat auki. Tämä on kriittinen vaihe, sillä proteiinit saattavat mennä sikinsokin ja voivat muodostaa komplekseja väärien proteiinien kanssa. Suojelevat esiliinaproteiinit pitävät ne sellaisessa tilassa, etteivät ne pääse laskostumaan väärin, huomauttaa Sistonen.
Syöpäsolussa on ilmeisesti laskostukseen erikoistuneita suojeluproteiineja jo valmiiksi paljon, jotta ne selviävät syöpäsolun kiihtyvästä aineenvaihdunnasta.
Geeniluenta selvittää hiljentyvät ja aktivoituvat geenit
Lea Sistonen tutkii geeniluentaa, jossa selvitetään, mitkä geenit hiljennetään ja mitkä aktivoituvat. Ainoastaan murto-osa kaikissa soluissa olevista samoista geeneistä aktivoituu tietyissä vaiheissa.
– Haasteita aiheuttaa muun muassa se, että geeniluenta yleensä hiljennetään silloin, kun solut alkavat tuottaa suojelevia proteiineja. Vain tietyt lämpösokkitekijät aktivoituvat. Estäjiä ja stimulaattoreita etsitään nyt kovasti. Tutkimme parhaillaan sitä, miten geeniluenta tapahtuu ja kuinka sitä pitäisi joskus estää syövässä. Jos soluissa on yliaktiivinen stressivaste, se pitää hiljentää, kertoo Lea Sistonen.
Kolikon toinen puoli on se, että esimerkiksi Alzheimerin taudissa ja Parkinsonin taudissa aivoihin kertyy ei-toiminnallisia proteiineja, jotka pitäisi poistaa, mutta jotka eivät jostakin syystä häviä. Siksi nämä proteiinit sakkautuvat aivoihin.
– Toisissa soluissa siis pitäisi edesauttaa stressinsietokykyä, kun taas joissakin soluissa sitä pitäisi estää. Nyt tutkitaankin eri lääkeaineita, joista toivotaan apua siihen, että syöpäsoluihin voitaisiin viedä estäjiä ja aivoihin stimulaattoreita, painottaa Sistonen.
LÄHDE: UllaToikkanen
http://www.laakarilehti.fi/uutinen.html?opcode=show/news_id=7515/news_db=web_lehti2006/type=1
tisdag 9 juni 2009
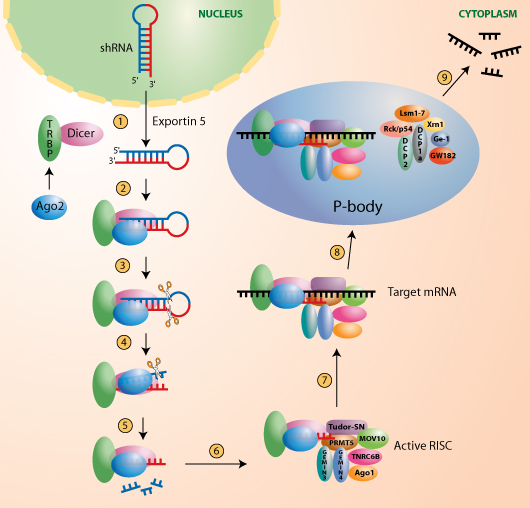
Ei- koodaava RNA, RNAi, Dicer , RISC
2009 muistiin
noncoding RNA,
RNAi,
Dicer,
TRBP,
PACT,
RISC,
miRNA, siRNA
microRNA, premicroRNA,
Katsaus 1.
MikroRNA-molekyylit ovat kehkeytyneet kahden vaiheen prosessilla, joka tuottavat noin 22 nukleotidin(nt) RNA molekyylejä. Ne säätelevät negatiivisesti (= vaimentavat) kohdegeeniensä ekspressiota post-transkriptionaalisella tasolla.
Primääriset miRNA molekyylit prosessoituvat prekursori miRNA:ksi ( pre-miRNA) mikroprosessorikompleksin välityksellä.
Kuva esittää miten tehdään sekvenssispesifinen geenin hiljentymä kohdegeenistä lähtevään mRNA-proteiiniin.
http://www.nature.com/nri/journal/v3/n11/images/nri1227-f1.jpg
Nämä pre-miRNA muodot pilkkoutuvat RNAasiIII Dicer entsyymillä ja niistä kehkeytyy kypsää miRNA muotoa, jotka ohjaavat RNA- indusoitua hiljentäjäkompleksia (RISC, RNA induced silencing Complex) kohdegeenistä kirjoitetulle lähetti-RNA:lle (mRNA) komplementaarisella sekvenssillä.
Tässä työssä tutkijat osoittavat, että Dicer-entsyymiä sisältävän kompleksin integraalinen komponentti on TRBP (HIV transactivating response RNA binding protein), mikä sisältää kolme kaksoissäikeistä RNA:ta sitovaa domeenia.
TRBP-sisältöisten kompleksien biokemiallinen analyysi paljastaa assosiaation Dicer-TRBP ja Argonaute-2( Ago-2) kesken.
http://www.invivogen.com/images/shRNA_pathway.jpg
Ago2 on RISC- kompleksin katalyyttinen moottori. Fysikaalinen assosiaatio näiden kesken varmistettiin Koeputkessa osoitettiin , että TRBP vaaditaan Ago2:n rekrytoimiseen siRNA:lle( small interfering RNA) , joka taas on sitoutunut Diceriin.
TRBP Knock out malli tekee Dicerin epästabiiliksi ja siitä seuraa, että miRNA biogeneesi jää tapahtumatta
Dicer-TRBP kompleksin vaje vähentää RISC-välitteistä reportterigeenin vaimenemista.
Nämä tulokset osoittavat, että Dicer-TRBP kompleksilla on osuutta miRNA prosessoinnin lisäksi myös RISC kompleksin kokoontumisen alustana.
LÄHDE1.
Chendrimada TP, Gregory RI et al. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. : Nature. 2005 Aug 4;436(7051):740-4. Epub 2005 Jun 22.
http://www.retrovirology.com/content/2/1/65
Katsaus 2.
Pienten RNA- molekyylien kautta tapahtuva geenien vaimeneminen näyttää olevan päätie eukaryoottisisten solujen säätelyssä. Tässä tiessä esiintyvien päätekijöiden identifioiminen on ollut intensiivisen tutkimuksen kohteena viime vuosina.
Ihmisellä pieniä RNA-molekyylejä tuottaa Dicer-entsyymi ja niitä kertyy vaikuttajakomplekseihin kutea RISC (RNA induced silencing complex) useine tekijöineen
joihin kuuluu hAgo2, mRNA:n kohteekseen valitseva endonukleaasi
ja TRBV (HIV-1 TAR RNA binding protein)
http://homepage.usask.ca/~vim458/advirol/SPCV/miRNA/figure5.jpg
ds RNA-sitova lisäproteiini, joka tunnetaan nimellä PACT (merkitsevä tekijä RNA-hiljennyksessä).
PACT liittyy 500kDA kompleksiin, joka sisältää seuraavat tekijät: Dicer, hAgo2 ja TRBP.
http://www1.qiagen.com/GeneGlobe/Pathways/tiny/Activation%20of%20NF-KappaB%20by%20PKR.jpg
http://www1.qiagen.com/GeneGlobe/PathwayView.aspx?pathwayID=14
(a protein activator of PKR = PACT)
Dicer-interaktio käsittää PACT:in kolmannen dsRNA:ta sitovan domaanin (dsRBD) ja Dicerin N-terminaalisen alueen, jossa on helikaasi-motiivi.
Kuten ei TRBP niin ei myös PACT ole tarpeen pre-mikroRNA:n pilkkoutumisreaktiossa.
Mutta kuitenkin PACT-tekijän puute vaikuttaa vahvasti kypsän miRNA:n kertymisiin in vivo ja kohtalaisesti vähentää siRNA:n tehoa indusoida RNA-interferenssiä.
Tämä tutkimus osoittaa, että ihmisen Dicer entsyymi (muista RNAasi III proteiinityypeistä poiketen) saattaa rekrytoida kaksi erilaista dsRBD-sisältävää proteiinia, jotka kiihdyttävät RISC- kompleksin muodostusta.
LÄHDE 2.
Yoontae Lee et al. The role of PACT in the RNA silencing pathway EMBO J. 2006 February 8; 25(3): 522–532. Published online 2006 January 19
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1383527
Muistiin RNA interferenssin osatekijöistä.
18/04/2008 21:51
|
noncoding RNA,
RNAi,
Dicer,
TRBP,
PACT,
RISC,
miRNA, siRNA
microRNA, premicroRNA,
Katsaus 1.
MikroRNA-molekyylit ovat kehkeytyneet kahden vaiheen prosessilla, joka tuottavat noin 22 nukleotidin(nt) RNA molekyylejä. Ne säätelevät negatiivisesti (= vaimentavat) kohdegeeniensä ekspressiota post-transkriptionaalisella tasolla.
Primääriset miRNA molekyylit prosessoituvat prekursori miRNA:ksi ( pre-miRNA) mikroprosessorikompleksin välityksellä.
Kuva esittää miten tehdään sekvenssispesifinen geenin hiljentymä kohdegeenistä lähtevään mRNA-proteiiniin.
http://www.nature.com/nri/journal/v3/n11/images/nri1227-f1.jpg
{kind=link}
Nämä pre-miRNA muodot pilkkoutuvat RNAasiIII Dicer entsyymillä ja niistä kehkeytyy kypsää miRNA muotoa, jotka ohjaavat RNA- indusoitua hiljentäjäkompleksia (RISC, RNA induced silencing Complex) kohdegeenistä kirjoitetulle lähetti-RNA:lle (mRNA) komplementaarisella sekvenssillä.
Tässä työssä tutkijat osoittavat, että Dicer-entsyymiä sisältävän kompleksin integraalinen komponentti on TRBP (HIV transactivating response RNA binding protein), mikä sisältää kolme kaksoissäikeistä RNA:ta sitovaa domeenia.
TRBP-sisältöisten kompleksien biokemiallinen analyysi paljastaa assosiaation Dicer-TRBP ja Argonaute-2( Ago-2) kesken.
http://www.invivogen.com/images/shRNA_pathway.jpg
{kind=link}
Ago2 on RISC- kompleksin katalyyttinen moottori. Fysikaalinen assosiaatio näiden kesken varmistettiin Koeputkessa osoitettiin , että TRBP vaaditaan Ago2:n rekrytoimiseen siRNA:lle( small interfering RNA) , joka taas on sitoutunut Diceriin.
TRBP Knock out malli tekee Dicerin epästabiiliksi ja siitä seuraa, että miRNA biogeneesi jää tapahtumatta
Dicer-TRBP kompleksin vaje vähentää RISC-välitteistä reportterigeenin vaimenemista.
Nämä tulokset osoittavat, että Dicer-TRBP kompleksilla on osuutta miRNA prosessoinnin lisäksi myös RISC kompleksin kokoontumisen alustana.
LÄHDE1.
Chendrimada TP, Gregory RI et al. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. : Nature. 2005 Aug 4;436(7051):740-4. Epub 2005 Jun 22.
http://www.retrovirology.com/content/2/1/65
Katsaus 2.
Pienten RNA- molekyylien kautta tapahtuva geenien vaimeneminen näyttää olevan päätie eukaryoottisisten solujen säätelyssä. Tässä tiessä esiintyvien päätekijöiden identifioiminen on ollut intensiivisen tutkimuksen kohteena viime vuosina.
Ihmisellä pieniä RNA-molekyylejä tuottaa Dicer-entsyymi ja niitä kertyy vaikuttajakomplekseihin kutea RISC (RNA induced silencing complex) useine tekijöineen
joihin kuuluu hAgo2, mRNA:n kohteekseen valitseva endonukleaasi
ja TRBV (HIV-1 TAR RNA binding protein)
http://homepage.usask.ca/~vim458/advirol/SPCV/miRNA/figure5.jpg
{kind=link}
ds RNA-sitova lisäproteiini, joka tunnetaan nimellä PACT (merkitsevä tekijä RNA-hiljennyksessä).
PACT liittyy 500kDA kompleksiin, joka sisältää seuraavat tekijät: Dicer, hAgo2 ja TRBP.
http://www1.qiagen.com/GeneGlobe/Pathways/tiny/Activation%20of%20NF-KappaB%20by%20PKR.jpg
{kind=link}
http://www1.qiagen.com/GeneGlobe/PathwayView.aspx?pathwayID=14
(a protein activator of PKR = PACT)
Dicer-interaktio käsittää PACT:in kolmannen dsRNA:ta sitovan domaanin (dsRBD) ja Dicerin N-terminaalisen alueen, jossa on helikaasi-motiivi.
Kuten ei TRBP niin ei myös PACT ole tarpeen pre-mikroRNA:n pilkkoutumisreaktiossa.
Mutta kuitenkin PACT-tekijän puute vaikuttaa vahvasti kypsän miRNA:n kertymisiin in vivo ja kohtalaisesti vähentää siRNA:n tehoa indusoida RNA-interferenssiä.
Tämä tutkimus osoittaa, että ihmisen Dicer entsyymi (muista RNAasi III proteiinityypeistä poiketen) saattaa rekrytoida kaksi erilaista dsRBD-sisältävää proteiinia, jotka kiihdyttävät RISC- kompleksin muodostusta.
LÄHDE 2.
Yoontae Lee et al. The role of PACT in the RNA silencing pathway EMBO J. 2006 February 8; 25(3): 522–532. Published online 2006 January 19
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1383527
Muistiin RNA interferenssin osatekijöistä.
18/04/2008 21:51
|
tisdag 19 maj 2009
Solusyklistä. Mitoosiin saapuminen ( A Lindqvistin mukaan)
LÄHDE: Lindqvist Arne. Regulation of CDK dephosphorylation in mitotic entry. Stockholm 2005. Karolinska Institutet. ISBN91-7140-362-0.
Abstraktin suomennosta, terminologiasta.
Näistä ovat mitoosin induktoreita tumassa Cdc25A (nukleaarinen; NLS ja NES segmentti)
ja mitoosin induktori tumassa sekä sytoplasmassa Cdc25B (nukleaarinen, NLS ja NES segmentti). Sen sijaan yksinään Cdc25C pysäyttää solun G2 vaiheeseen.
http://www.nature.com/nrc/journal/v4/n5/images/nrc1361-f3.jpg
Solusyklin progressio vaatii sykliinistä (cyclin, Cyc) riippuvien kinaasien (Cyclin dependent kinase, Cdk) kaskadin, jossa kompleksin keskenään tehneet kinaasit ja sykliinit kompleksin aktivoiduttua ovat moottoreina.
http://nobelprize.org/nobel_prizes/medicine/laureates/2001/press-sv.html
G2-faasissa sykliini B1/Cdk 1 pidetään inaktiivina siten, että Cdk1 – molekyylin aktiivikohdassa on fosforylaatio.
Ihmisen sykliini / Cdk – komplekseja pystyy aktivoimaan kolme Cdc25- defosforylaasi- perheen jäsentä, jotka ovat kaksoisspesifisyyttä omaavia fosfataaseja. Ne ovat Cdc25A, Cdc25B ja Cdc25C. Tutkijaryhmä pyrki saamaan tarkempaa tietoa sykliini B1/ Cdk 1 aktivaation säätelystä mitoosiin saapumisvaiheessa.
Työhypoteesina oli se, että solu kontrolloi osittain sykliini B1/ Cdk1- aktivaatiota säätelemällä Cdc25- proteiinien asettumista solussa. Tämän takia tutkittiin Cdc25A ja Cdc25B- molekyylien intrasellulaarista sijaintia. Havaittiin, että molemmat (Cdc25A ja Cdc25B) sijaitsevat nukleaarisesti, tumassa, vaikka ne jatkuvasti sukkuloivat sytoplasman ja tuman väliä ( shuttle) .
Jos soluja käsiteltiin Cdc25A- tai Cdc25B-vähentävästi (siRNA avulla) siten, että Cdc25C-vaikutukseen ei puututtu, aiheutui viivästystä mitoosiin saapumisessa.
Mutta jos sekä Cdc25A että Cdc25B vähenivät ja jäljellä oli vain Cdc25C-vaikutus, solu pysähtyi G2-vaiheeseensa.
Seurattaessa sykliini B1/Cdk 1- aktivaatiota fosfospesifisillä antibodeilla, havaittiin aktiivia kompleksia sykliini B1/Cdk 1 kertyvän ensiksi centrosomeihin hieman ennen kuin ne alkavat migroitua erilleen. Sitten aktivaatio leviää sytoplasmaan ja kun 50 % komplekseista on aktivoituneena, sykliini B1/Cdk 1 sytoplasmastakin alkaa translokoituu tumaan. Translokaation aikana sykliini B1/Cdk 1- kompleksin aktivaatio edistyy samalla tavalla tumassa kuin sytoplasman puolella. Tutkimuksessa ei havaittu mitään sykliini B1/Cdk 1-inaktivaatiota myöhäisessä metafaasissa. Saatu tieto tukee käsitystä bistabiliteetistä, joka hallitseee sykliini B1/Cdk 1 aktivaatiota.”
Terminologiasta muutama seikka:
MITOOTTINEN JAKAANTUMINEN tuottaa kaksi samanlaista tytärsolua.
Kirjan tekstiin kuuluu kuva mitoosiin saapumisvaiheesta, ja tässä on seuraavat spekulatiiviset komponentit esitettynä: vain tekstissä mainittuja molekyylejä käytetään kaavassa. (Cdc25fosfataasit eli defosforylaasit ja sykliini/Cdk-kompleksit, Cyc /Cdk eli solumoottorit, kinaasit ).
Aikajana: G2 ;PROFAASI; PROMETAFAASI; METAFAASI.
(Tapahtumat sytoplasmassa.
Tapahtumat sytoplasman ja tuman välisissä kuljetuksissa (shuttle)
Tapahtumat tumassa)
Kun riittävä määrä sykliini B1/Cdk 1 kompleksia alkoi olla centrosomeissa, määrä aktivoitui Cdc25B- tekijällä-jarruttavan fosfaatin poistamisella.
Centrosomaalinen sykliiniB1/Cdk 1 aktivaatio johti centrosomien migraatioon.
Tuman sykliini A/Cdk2 aktivoitui sekä Cdc25A että Cdc25B- tekijöitten avulla.
Sykliini A /Cdk 2 ja sykliini B1/Cdk 1 voivat aktivoitua synkronisoidusti, koska sekä Cdc25- ryhmä ja sykliini/Cdk-kompleksit voivat sukkuloida ( shuttle) sytoplasman ja tuman väliä.
Tuman sykliiniA1/Cdk 2 aktivoituminen johti DNA-kondensaation alkamiseen.
Aktivoitunutta sykliini B1/Cdk 1 translokoitui tumaan kiihdyttämään DNA-kondensaatiota.
Kun tumakalvo oli hajonnut, Cdc25C piti yllä maksimaalista sykliini B1/Cdk 1-aktiviteettia.
Mahdollisesti Cdc25A ja Cdc25B myös jollain tavalla osallistuvat.
Cdc25C siis huolehti propagoitumisen, vaikka ei aktivoitumista sykliini B1/Cdk1-komponentissa.
bhs.smuhsd.org/…/Cell_Replication.html
Tulevia tutkimuskohteita mainittiin
Moni mitoosiin saapumisen mekanismiin liittyvä kysymys on vielä ratkaisematta Todennäköisimmin tähän osallistuu vielä tunnistamattomia valkuaisaineita. Tutkija mainitsee muutamia kiintoisia tutkimuskohteita kuten
Millä tavalla, missä ja milloin Cdc25B hajoaa? On mahdollista, että Cdc25B ohjataan hajoitettavaksi SCF-avulla (Skp/Cullin/F-box protein). Jos niin on, mikä F-box välittää interaktion?
Onko Pin 1 osallistumassa Cdc25B-aktivaatioon?
http://www.nature.com/nrm/journal/v8/n11/images/nrm2261-f3.jpg
Jos se osallistuu, kilpaileeko se pelkästään 14-3-3-kohdasta Cdc25B:n kanssa vai onko lisämekanismeja? ( Kaikki 14-3-3 proteiinit sitovat fosforyloituja epitooppeja. Kaikki Cdc25 voivat sitoutua 14-3-3- proteiineihin, jotka säätelevät monia soluprosesseja).
Stimuloiko Plk 1( Polo-like kinase 1), Aurora A-, CyclinA/Cdk2 -tai Cyclin B1/Cdk 1-fosforylaatio Pin 1 sitoutumista? Tai päinvastoin: kiihdyttääkö Pin 1 isomerisaatio niiden fosforyloitumista Cdc25B:llä? Missä ja milloin Pin 1 on interaktiossa Cdc25B:n kanssa? (Pin 1 on isomeraasi, joka voi muuttaa kohteen struktuuria. Pin1 tunnistaa Cyc/Cdk-kompleksin).
Mikä on Cdc25C:n osuus mitoosissa? Interferenssitutkimuksilla on osoitettu, että siRNA tätä Cdc25C.tekijää kohtaan tuottaa viivästyneen myöhäisprometafaasin. Onko sykliini B1/Cdk 1 aktiivisuus hieman alempi näissä soluissa ? Esim. muuttaako Cdc25C –puute bistabiliteettiominaisuuksia sykliini B1/Cdk 1 aktiviteetissa? Jos niin on, onko niissä soluissa mikrotubulaarisia dynamiikan eroja?
Onko sentromeerisukkulajärjestelmän kontrollikohta aktivoitunut ? ( spindle assembly checkpoint – tässä katsotaan olevan p38-riippuvainen säätö).
Milloin, miten ja missä sykliiniA/Cdk2 aktivoituu ? Toistaiseksi(2005) on tehty vain in vitro tutkimuksia tämän kompleksin aktivoitumisen suhteen. Heijastaako in vitro-tutkimukset sykliini A/Cdk2-aktiivisuutta in vivo-olosuhteissa?
Käännös 2005-11-08 18:53
Päivitys 2008-04-01 00:26. Päivitys 24.6. 2009, lisään lähdekuvia netistä.
Tämä väitöskirja antaa hyvän yleiskatsauksen tähän asti tunnetuista solysyklikäsityksistä ja availee näkymiä seuraaviin tieteellisiin solusyklisondeerauksiin .
Otan näitä tekijöitä tähän kotisivulle talteen Cell Cycle otsikkon alle, jos tässä tulee vielä pohdittavaksi strategioita jonkin pandemian tms solusyklimodifikaation takia.
Toistan vielä abstraktin ja otan sen 19.11. 2016 uudelle sivulle. Löydän siihen myös nettilinkin KI:sta. .
Abstraktin suomennosta, terminologiasta.
- sykliini B1/Cdk 1- Cdc25 fosfataasit eli defosforylaasit:
Näistä ovat mitoosin induktoreita tumassa Cdc25A (nukleaarinen; NLS ja NES segmentti)
ja mitoosin induktori tumassa sekä sytoplasmassa Cdc25B (nukleaarinen, NLS ja NES segmentti). Sen sijaan yksinään Cdc25C pysäyttää solun G2 vaiheeseen.
http://www.nature.com/nrc/journal/v4/n5/images/nrc1361-f3.jpg
{kind=link}
- Exportiini-1 ( inhibiittori)
Solusyklin progressio vaatii sykliinistä (cyclin, Cyc) riippuvien kinaasien (Cyclin dependent kinase, Cdk) kaskadin, jossa kompleksin keskenään tehneet kinaasit ja sykliinit kompleksin aktivoiduttua ovat moottoreina.
http://nobelprize.org/nobel_prizes/medicine/laureates/2001/press-sv.html
- Varhaisten mitoottisten tapahtumien avainsäätelijä on sykliini B1/Cdk 1-kompleksi.
G2-faasissa sykliini B1/Cdk 1 pidetään inaktiivina siten, että Cdk1 – molekyylin aktiivikohdassa on fosforylaatio.
- Mikä on Cdc25 fosfataasien ( defosforylaasien) osuus?
Ihmisen sykliini / Cdk – komplekseja pystyy aktivoimaan kolme Cdc25- defosforylaasi- perheen jäsentä, jotka ovat kaksoisspesifisyyttä omaavia fosfataaseja. Ne ovat Cdc25A, Cdc25B ja Cdc25C. Tutkijaryhmä pyrki saamaan tarkempaa tietoa sykliini B1/ Cdk 1 aktivaation säätelystä mitoosiin saapumisvaiheessa.
Työhypoteesina oli se, että solu kontrolloi osittain sykliini B1/ Cdk1- aktivaatiota säätelemällä Cdc25- proteiinien asettumista solussa. Tämän takia tutkittiin Cdc25A ja Cdc25B- molekyylien intrasellulaarista sijaintia. Havaittiin, että molemmat (Cdc25A ja Cdc25B) sijaitsevat nukleaarisesti, tumassa, vaikka ne jatkuvasti sukkuloivat sytoplasman ja tuman väliä ( shuttle) .
- NES
- NLS
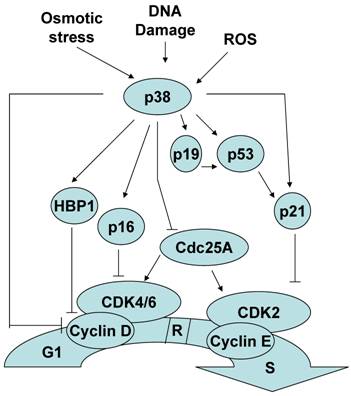
- Ultravioletille (UV) valolle reagoi Cdc25B translokoitumalla osittain tuman puolelle. Tähän siirtymään, translokaatioon, vaadittiin Cdc25B NES, p38 MAPK aktiviteetti ja vapaa14-3-3- sitoutuminen.
{kind=link}
- Ehdotettiin seuraavaa: stressivasteena p38 MAPK välitti 14-3-3-sitoutumista Cdc25B-molekyyliin, mikä johti sytoplasmiseen kertymään. (Jokainen Cdc25 pystyy sitomaan 14-3-3 proteiinia, jolloin Cdc25 inhiboituu) .
- RNA-interferenssimetodia käyttämällä (RNAi) vähennettiin eri Cdc25-molekyylien osuuksia, jotta saataisiin esiin niitten yksilölliset tehtävät.
Jos soluja käsiteltiin Cdc25A- tai Cdc25B-vähentävästi (siRNA avulla) siten, että Cdc25C-vaikutukseen ei puututtu, aiheutui viivästystä mitoosiin saapumisessa.
Mutta jos sekä Cdc25A että Cdc25B vähenivät ja jäljellä oli vain Cdc25C-vaikutus, solu pysähtyi G2-vaiheeseensa.
- Tästä pääätellään, että mitoosin induktorit ovat Cdc25A ja Cdc25B. Ajallisesti seurattaessa tapahtumaa ( time-lapse movies), vaikuttaa siltä, että Cdc25A säätää pääasiassa tuman tapahtumia, kun taas Cdc25B mahdollisesti osallistuu sekä tuman että sytoplasman tapahtumien säätelyyn.
{kind=link}
- Jos RNA-interfererenssimetodilla (RNAi ja siRNA) vähennettiin Cdc25B, mutta ei Cdc25A-molekyyliä, seurasi korkeampia pitoisuuksia fosforyloitua sykliini B1/Cdk 1 kompleksia erkanemattomissa sentrosomeissa kontrollisoluihin verrattuna. Tästä voitiin päätellä, että CdcB25 osallistui prosessiin, mistä seuraa aktiivin sykliini B1/Cdk 1- kompleksin akkumuloitumista solun centrosomeihin.
Seurattaessa sykliini B1/Cdk 1- aktivaatiota fosfospesifisillä antibodeilla, havaittiin aktiivia kompleksia sykliini B1/Cdk 1 kertyvän ensiksi centrosomeihin hieman ennen kuin ne alkavat migroitua erilleen. Sitten aktivaatio leviää sytoplasmaan ja kun 50 % komplekseista on aktivoituneena, sykliini B1/Cdk 1 sytoplasmastakin alkaa translokoituu tumaan. Translokaation aikana sykliini B1/Cdk 1- kompleksin aktivaatio edistyy samalla tavalla tumassa kuin sytoplasman puolella. Tutkimuksessa ei havaittu mitään sykliini B1/Cdk 1-inaktivaatiota myöhäisessä metafaasissa. Saatu tieto tukee käsitystä bistabiliteetistä, joka hallitseee sykliini B1/Cdk 1 aktivaatiota.”
Terminologiasta muutama seikka:
- P38 MAPK tunnetaan G1/S-siirtymän säätelijänä rb avulla. P38 säätelee p53 ja siirtymää faasirajalla G1/S.
MITOOTTINEN JAKAANTUMINEN tuottaa kaksi samanlaista tytärsolua.
Kirjan tekstiin kuuluu kuva mitoosiin saapumisvaiheesta, ja tässä on seuraavat spekulatiiviset komponentit esitettynä: vain tekstissä mainittuja molekyylejä käytetään kaavassa. (Cdc25fosfataasit eli defosforylaasit ja sykliini/Cdk-kompleksit, Cyc /Cdk eli solumoottorit, kinaasit ).
Aikajana: G2 ;PROFAASI; PROMETAFAASI; METAFAASI.
(Tapahtumat sytoplasmassa.
Tapahtumat sytoplasman ja tuman välisissä kuljetuksissa (shuttle)
Tapahtumat tumassa)
- G2 vaiheessa
Kun riittävä määrä sykliini B1/Cdk 1 kompleksia alkoi olla centrosomeissa, määrä aktivoitui Cdc25B- tekijällä-jarruttavan fosfaatin poistamisella.
Centrosomaalinen sykliiniB1/Cdk 1 aktivaatio johti centrosomien migraatioon.
Tuman sykliini A/Cdk2 aktivoitui sekä Cdc25A että Cdc25B- tekijöitten avulla.
Sykliini A /Cdk 2 ja sykliini B1/Cdk 1 voivat aktivoitua synkronisoidusti, koska sekä Cdc25- ryhmä ja sykliini/Cdk-kompleksit voivat sukkuloida ( shuttle) sytoplasman ja tuman väliä.
- PROFAASI
{kind=link}
Tuman sykliiniA1/Cdk 2 aktivoituminen johti DNA-kondensaation alkamiseen.
- PROMETAFAASI
{kind=link}
Aktivoitunutta sykliini B1/Cdk 1 translokoitui tumaan kiihdyttämään DNA-kondensaatiota.
Kun tumakalvo oli hajonnut, Cdc25C piti yllä maksimaalista sykliini B1/Cdk 1-aktiviteettia.
Mahdollisesti Cdc25A ja Cdc25B myös jollain tavalla osallistuvat.
Cdc25C siis huolehti propagoitumisen, vaikka ei aktivoitumista sykliini B1/Cdk1-komponentissa.
- METAFAASI
bhs.smuhsd.org/…/Cell_Replication.html
Tulevia tutkimuskohteita mainittiin
Moni mitoosiin saapumisen mekanismiin liittyvä kysymys on vielä ratkaisematta Todennäköisimmin tähän osallistuu vielä tunnistamattomia valkuaisaineita. Tutkija mainitsee muutamia kiintoisia tutkimuskohteita kuten
Millä tavalla, missä ja milloin Cdc25B hajoaa? On mahdollista, että Cdc25B ohjataan hajoitettavaksi SCF-avulla (Skp/Cullin/F-box protein). Jos niin on, mikä F-box välittää interaktion?
Onko Pin 1 osallistumassa Cdc25B-aktivaatioon?
http://www.nature.com/nrm/journal/v8/n11/images/nrm2261-f3.jpg
{kind=link}
Jos se osallistuu, kilpaileeko se pelkästään 14-3-3-kohdasta Cdc25B:n kanssa vai onko lisämekanismeja? ( Kaikki 14-3-3 proteiinit sitovat fosforyloituja epitooppeja. Kaikki Cdc25 voivat sitoutua 14-3-3- proteiineihin, jotka säätelevät monia soluprosesseja).
Stimuloiko Plk 1( Polo-like kinase 1), Aurora A-, CyclinA/Cdk2 -tai Cyclin B1/Cdk 1-fosforylaatio Pin 1 sitoutumista? Tai päinvastoin: kiihdyttääkö Pin 1 isomerisaatio niiden fosforyloitumista Cdc25B:llä? Missä ja milloin Pin 1 on interaktiossa Cdc25B:n kanssa? (Pin 1 on isomeraasi, joka voi muuttaa kohteen struktuuria. Pin1 tunnistaa Cyc/Cdk-kompleksin).
- KUVA: Polo-like kinase
{kind=link}
- KUVA: Aurora A, B
{kind=link}
Mikä on Cdc25C:n osuus mitoosissa? Interferenssitutkimuksilla on osoitettu, että siRNA tätä Cdc25C.tekijää kohtaan tuottaa viivästyneen myöhäisprometafaasin. Onko sykliini B1/Cdk 1 aktiivisuus hieman alempi näissä soluissa ? Esim. muuttaako Cdc25C –puute bistabiliteettiominaisuuksia sykliini B1/Cdk 1 aktiviteetissa? Jos niin on, onko niissä soluissa mikrotubulaarisia dynamiikan eroja?
Onko sentromeerisukkulajärjestelmän kontrollikohta aktivoitunut ? ( spindle assembly checkpoint – tässä katsotaan olevan p38-riippuvainen säätö).
- KUVA Spindle assembly checkpoint
Milloin, miten ja missä sykliiniA/Cdk2 aktivoituu ? Toistaiseksi(2005) on tehty vain in vitro tutkimuksia tämän kompleksin aktivoitumisen suhteen. Heijastaako in vitro-tutkimukset sykliini A/Cdk2-aktiivisuutta in vivo-olosuhteissa?
Miten solusyklin faasi vaikuttaa paikallistumisen dynamiikkaan ( localisation dynamics)? Lokalisaatiosignaaleihin sekä proteiinien ja rakenteitten interaktioihin mahdollisesti vaikuttavat erilaiset expressiotasot ja posttranslationaaliset modifikaatiot. Nykyinen edistys FRAP-analyyseissä ( Fluorescence Recovery After Photobleaching) mahdollistanee lähitulevaisuudessa multippeleitten interaktiokompleksien kokojen ja diffuusiovakioitten arvioimisen (Sprague et McNally 2005). Kun tehdään systemaattisia FRAP ja FLIP tutkimuksia useista solukohdista eri solusyklivaiheitten aikana lukuisilla solusyklin säätelijöillä pystyttäisiin tekemään johtopäätöksiä interaktiokompleksien koosta ja liikkuuvuudesta.
Käännös 2005-11-08 18:53
Päivitys 2008-04-01 00:26. Päivitys 24.6. 2009, lisään lähdekuvia netistä.
Tämä väitöskirja antaa hyvän yleiskatsauksen tähän asti tunnetuista solysyklikäsityksistä ja availee näkymiä seuraaviin tieteellisiin solusyklisondeerauksiin .
Otan näitä tekijöitä tähän kotisivulle talteen Cell Cycle otsikkon alle, jos tässä tulee vielä pohdittavaksi strategioita jonkin pandemian tms solusyklimodifikaation takia.
Toistan vielä abstraktin ja otan sen 19.11. 2016 uudelle sivulle. Löydän siihen myös nettilinkin KI:sta. .
Prenumerera på:
Kommentarer (Atom)