
Abstraktin suomennosta, terminologiasta.
- sykliini B1/Cdk 1- Cdc25 fosfataasit eli defosforylaasit:
Näistä ovat mitoosin induktoreita tumassa Cdc25A (nukleaarinen; NLS ja NES segmentti)
ja mitoosin induktori tumassa sekä sytoplasmassa Cdc25B (nukleaarinen, NLS ja NES segmentti). Sen sijaan yksinään Cdc25C pysäyttää solun G2 vaiheeseen.
http://www.nature.com/nrc/journal/v4/n5/images/nrc1361-f3.jpg
{kind=link}
- Exportiini-1 ( inhibiittori)
Solusyklin progressio vaatii sykliinistä (cyclin, Cyc) riippuvien kinaasien (Cyclin dependent kinase, Cdk) kaskadin, jossa kompleksin keskenään tehneet kinaasit ja sykliinit kompleksin aktivoiduttua ovat moottoreina.
http://nobelprize.org/nobel_prizes/medicine/laureates/2001/press-sv.html
- Varhaisten mitoottisten tapahtumien avainsäätelijä on sykliini B1/Cdk 1-kompleksi.
G2-faasissa sykliini B1/Cdk 1 pidetään inaktiivina siten, että Cdk1 – molekyylin aktiivikohdassa on fosforylaatio.
- Mikä on Cdc25 fosfataasien ( defosforylaasien) osuus?
Ihmisen sykliini / Cdk – komplekseja pystyy aktivoimaan kolme Cdc25- defosforylaasi- perheen jäsentä, jotka ovat kaksoisspesifisyyttä omaavia fosfataaseja. Ne ovat Cdc25A, Cdc25B ja Cdc25C. Tutkijaryhmä pyrki saamaan tarkempaa tietoa sykliini B1/ Cdk 1 aktivaation säätelystä mitoosiin saapumisvaiheessa.
Työhypoteesina oli se, että solu kontrolloi osittain sykliini B1/ Cdk1- aktivaatiota säätelemällä Cdc25- proteiinien asettumista solussa. Tämän takia tutkittiin Cdc25A ja Cdc25B- molekyylien intrasellulaarista sijaintia. Havaittiin, että molemmat (Cdc25A ja Cdc25B) sijaitsevat nukleaarisesti, tumassa, vaikka ne jatkuvasti sukkuloivat sytoplasman ja tuman väliä ( shuttle) .
- NES
- NLS
- Ultravioletille (UV) valolle reagoi Cdc25B translokoitumalla osittain tuman puolelle. Tähän siirtymään, translokaatioon, vaadittiin Cdc25B NES, p38 MAPK aktiviteetti ja vapaa14-3-3- sitoutuminen.
{kind=link}
- Ehdotettiin seuraavaa: stressivasteena p38 MAPK välitti 14-3-3-sitoutumista Cdc25B-molekyyliin, mikä johti sytoplasmiseen kertymään. (Jokainen Cdc25 pystyy sitomaan 14-3-3 proteiinia, jolloin Cdc25 inhiboituu) .
- RNA-interferenssimetodia käyttämällä (RNAi) vähennettiin eri Cdc25-molekyylien osuuksia, jotta saataisiin esiin niitten yksilölliset tehtävät.
Jos soluja käsiteltiin Cdc25A- tai Cdc25B-vähentävästi (siRNA avulla) siten, että Cdc25C-vaikutukseen ei puututtu, aiheutui viivästystä mitoosiin saapumisessa.
Mutta jos sekä Cdc25A että Cdc25B vähenivät ja jäljellä oli vain Cdc25C-vaikutus, solu pysähtyi G2-vaiheeseensa.
- Tästä pääätellään, että mitoosin induktorit ovat Cdc25A ja Cdc25B. Ajallisesti seurattaessa tapahtumaa ( time-lapse movies), vaikuttaa siltä, että Cdc25A säätää pääasiassa tuman tapahtumia, kun taas Cdc25B mahdollisesti osallistuu sekä tuman että sytoplasman tapahtumien säätelyyn.
{kind=link}
- Jos RNA-interfererenssimetodilla (RNAi ja siRNA) vähennettiin Cdc25B, mutta ei Cdc25A-molekyyliä, seurasi korkeampia pitoisuuksia fosforyloitua sykliini B1/Cdk 1 kompleksia erkanemattomissa sentrosomeissa kontrollisoluihin verrattuna. Tästä voitiin päätellä, että CdcB25 osallistui prosessiin, mistä seuraa aktiivin sykliini B1/Cdk 1- kompleksin akkumuloitumista solun centrosomeihin.
Seurattaessa sykliini B1/Cdk 1- aktivaatiota fosfospesifisillä antibodeilla, havaittiin aktiivia kompleksia sykliini B1/Cdk 1 kertyvän ensiksi centrosomeihin hieman ennen kuin ne alkavat migroitua erilleen. Sitten aktivaatio leviää sytoplasmaan ja kun 50 % komplekseista on aktivoituneena, sykliini B1/Cdk 1 sytoplasmastakin alkaa translokoituu tumaan. Translokaation aikana sykliini B1/Cdk 1- kompleksin aktivaatio edistyy samalla tavalla tumassa kuin sytoplasman puolella. Tutkimuksessa ei havaittu mitään sykliini B1/Cdk 1-inaktivaatiota myöhäisessä metafaasissa. Saatu tieto tukee käsitystä bistabiliteetistä, joka hallitseee sykliini B1/Cdk 1 aktivaatiota.”
Terminologiasta muutama seikka:
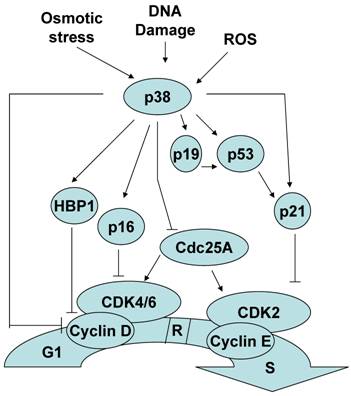
- P38 MAPK tunnetaan G1/S-siirtymän säätelijänä rb avulla. P38 säätelee p53 ja siirtymää faasirajalla G1/S.
MITOOTTINEN JAKAANTUMINEN tuottaa kaksi samanlaista tytärsolua.
Kirjan tekstiin kuuluu kuva mitoosiin saapumisvaiheesta, ja tässä on seuraavat spekulatiiviset komponentit esitettynä: vain tekstissä mainittuja molekyylejä käytetään kaavassa. (Cdc25fosfataasit eli defosforylaasit ja sykliini/Cdk-kompleksit, Cyc /Cdk eli solumoottorit, kinaasit ).
Aikajana: G2 ;PROFAASI; PROMETAFAASI; METAFAASI.
(Tapahtumat sytoplasmassa.
Tapahtumat sytoplasman ja tuman välisissä kuljetuksissa (shuttle)
Tapahtumat tumassa)
- G2 vaiheessa
Kun riittävä määrä sykliini B1/Cdk 1 kompleksia alkoi olla centrosomeissa, määrä aktivoitui Cdc25B- tekijällä-jarruttavan fosfaatin poistamisella.
Centrosomaalinen sykliiniB1/Cdk 1 aktivaatio johti centrosomien migraatioon.
Tuman sykliini A/Cdk2 aktivoitui sekä Cdc25A että Cdc25B- tekijöitten avulla.
Sykliini A /Cdk 2 ja sykliini B1/Cdk 1 voivat aktivoitua synkronisoidusti, koska sekä Cdc25- ryhmä ja sykliini/Cdk-kompleksit voivat sukkuloida ( shuttle) sytoplasman ja tuman väliä.
- PROFAASI
{kind=link}
Tuman sykliiniA1/Cdk 2 aktivoituminen johti DNA-kondensaation alkamiseen.
- PROMETAFAASI
{kind=link}
Aktivoitunutta sykliini B1/Cdk 1 translokoitui tumaan kiihdyttämään DNA-kondensaatiota.
Kun tumakalvo oli hajonnut, Cdc25C piti yllä maksimaalista sykliini B1/Cdk 1-aktiviteettia.
Mahdollisesti Cdc25A ja Cdc25B myös jollain tavalla osallistuvat.
Cdc25C siis huolehti propagoitumisen, vaikka ei aktivoitumista sykliini B1/Cdk1-komponentissa.
- METAFAASI
bhs.smuhsd.org/…/Cell_Replication.html
Tulevia tutkimuskohteita mainittiin
Moni mitoosiin saapumisen mekanismiin liittyvä kysymys on vielä ratkaisematta Todennäköisimmin tähän osallistuu vielä tunnistamattomia valkuaisaineita. Tutkija mainitsee muutamia kiintoisia tutkimuskohteita kuten
Millä tavalla, missä ja milloin Cdc25B hajoaa? On mahdollista, että Cdc25B ohjataan hajoitettavaksi SCF-avulla (Skp/Cullin/F-box protein). Jos niin on, mikä F-box välittää interaktion?
Onko Pin 1 osallistumassa Cdc25B-aktivaatioon?
http://www.nature.com/nrm/journal/v8/n11/images/nrm2261-f3.jpg
{kind=link}
Jos se osallistuu, kilpaileeko se pelkästään 14-3-3-kohdasta Cdc25B:n kanssa vai onko lisämekanismeja? ( Kaikki 14-3-3 proteiinit sitovat fosforyloituja epitooppeja. Kaikki Cdc25 voivat sitoutua 14-3-3- proteiineihin, jotka säätelevät monia soluprosesseja).
Stimuloiko Plk 1( Polo-like kinase 1), Aurora A-, CyclinA/Cdk2 -tai Cyclin B1/Cdk 1-fosforylaatio Pin 1 sitoutumista? Tai päinvastoin: kiihdyttääkö Pin 1 isomerisaatio niiden fosforyloitumista Cdc25B:llä? Missä ja milloin Pin 1 on interaktiossa Cdc25B:n kanssa? (Pin 1 on isomeraasi, joka voi muuttaa kohteen struktuuria. Pin1 tunnistaa Cyc/Cdk-kompleksin).
- KUVA: Polo-like kinase
{kind=link}
- KUVA: Aurora A, B
{kind=link}
Mikä on Cdc25C:n osuus mitoosissa? Interferenssitutkimuksilla on osoitettu, että siRNA tätä Cdc25C.tekijää kohtaan tuottaa viivästyneen myöhäisprometafaasin. Onko sykliini B1/Cdk 1 aktiivisuus hieman alempi näissä soluissa ? Esim. muuttaako Cdc25C –puute bistabiliteettiominaisuuksia sykliini B1/Cdk 1 aktiviteetissa? Jos niin on, onko niissä soluissa mikrotubulaarisia dynamiikan eroja?
Onko sentromeerisukkulajärjestelmän kontrollikohta aktivoitunut ? ( spindle assembly checkpoint – tässä katsotaan olevan p38-riippuvainen säätö).
- KUVA Spindle assembly checkpoint
Milloin, miten ja missä sykliiniA/Cdk2 aktivoituu ? Toistaiseksi(2005) on tehty vain in vitro tutkimuksia tämän kompleksin aktivoitumisen suhteen. Heijastaako in vitro-tutkimukset sykliini A/Cdk2-aktiivisuutta in vivo-olosuhteissa?
Miten solusyklin faasi vaikuttaa paikallistumisen dynamiikkaan ( localisation dynamics)? Lokalisaatiosignaaleihin sekä proteiinien ja rakenteitten interaktioihin mahdollisesti vaikuttavat erilaiset expressiotasot ja posttranslationaaliset modifikaatiot. Nykyinen edistys FRAP-analyyseissä ( Fluorescence Recovery After Photobleaching) mahdollistanee lähitulevaisuudessa multippeleitten interaktiokompleksien kokojen ja diffuusiovakioitten arvioimisen (Sprague et McNally 2005). Kun tehdään systemaattisia FRAP ja FLIP tutkimuksia useista solukohdista eri solusyklivaiheitten aikana lukuisilla solusyklin säätelijöillä pystyttäisiin tekemään johtopäätöksiä interaktiokompleksien koosta ja liikkuuvuudesta.
Käännös 2005-11-08 18:53
Päivitys 2008-04-01 00:26. Päivitys 24.6. 2009, lisään lähdekuvia netistä.
Tämä väitöskirja antaa hyvän yleiskatsauksen tähän asti tunnetuista solysyklikäsityksistä ja availee näkymiä seuraaviin tieteellisiin solusyklisondeerauksiin .
Otan näitä tekijöitä tähän kotisivulle talteen Cell Cycle otsikkon alle, jos tässä tulee vielä pohdittavaksi strategioita jonkin pandemian tms solusyklimodifikaation takia.
Toistan vielä abstraktin ja otan sen 19.11. 2016 uudelle sivulle. Löydän siihen myös nettilinkin KI:sta. .
Inga kommentarer:
Skicka en kommentar