Respondentti: Tri Nidal Chosheh.
Väitösaihe: Methylome and Transcriptome Profiling of Hepatocytes Derived from Human Pluripotenti Stem Cells.
http://hdl.handle.net/2077/54951

lördag 24 februari 2018
Thesis ubikitiini-proteosomi järjestelmästä kliinisissä taudeissa diagnostiikassa
Simon Sjödin. Evaluation of the Endo-Lysosomal System and the Ubiquitin-Proteosome System in Neurodegenerative Diseases.
pääpainotus oli diagnostiikaqssa ja biomarköörienlöytämisessä, eikävarinaisesti ubikitiiniproteosomijärjestelmässä, muta otan sen mitä löydän tästä jälkimmäisestä asiasta.
Fakultetsopponent Janne Lehtiö, professor KI STH.
http://hdl.handle.net/2077/54533
Suomennosta abstraktin sisällöstä:
Neurodegeneraatio on prosessi, jossa neuronisoluja katoaa ja oireista voidaan päätellä, miltä keskushermostoalueelta oireet johtuvat. Neurodegeneratiivisiin tauteihin lasketaan AD (Alzheimerin tauti) , PD (Parkinsonin tauti) ja FTD (frontotemporaalinen dementia) ja niissä kaikissa kertyy proteiiniaggrekaatteja, mikä viittaa näiden tuotannon tai vaihtumisen/hajottamisen olevan epäkunnossa. Täten toimimaton proteostaasi (proteiinien epätasapaino) näyttäisi olevan kyseessä neurodegeneratiivisissa taudeissa.
AD- taudissa on vähentynyt endosyyttisten ja autofagisten rakkuloiden vaihtuvuus ja niin kertyy aivokudokseen endolysosomaalisia valkuaisaineita ja käyttymätöntä ubikitiiniä.
PD- taudissa on viitettä lysosomien vikatoiminnasta tautiriskien ja geneettisten muuntumisten liittyessä lysosomaalisiin kertymishäiriöihin ja taudin vikuuttamien alueitten lysosomaalisten proteiinien vähentyneestä esiintymästä.
FTD:ssä tautia aiheuttavat mutaatiot ja geneettiset riskitekijät viittaavat autofagisen ja endosomaalisen järjestelmän muuntuneen toiminnan olevan osallisena patogeneesissä.
Väitöstyö selvittää proteostaasin vikatoimintaa aivoselkäydinnesteestä käsin- ja tähän kehitetään mittausmenetelmä. Siinä mitataan autofagisen ja endolysosomaalisen systeemin ja ubikitiini-proteosomi-systeemin proteiineja.
1) LAMP2, lysosomaali kalvoproteiini
2) kokopitkä ubikitiini ( joka on 9 ubikitiini-monomeerin pituinen)
3) endolysosomaalisten proteiinien paneli
Kehitetyllä menetelmällä analysoitiin mainittujen tautien (myös FTD alatyyppien ja FTD mutaation kantajien) likvornäytteet.
Johtopäätöksiä mm.
Mahdollisia biomerkitsijöinä voisi toimia likvorin ubikitiini AD-taudissa (korkea) ja endolysosomaaliset proteiinit ja ubikitiini PD-taudissa (matalat) kontrolleihin verrattuna. FTD taudissa löydöt dysfunktionaalisista proteiineista eivät olleet välittömästi havaittavissa.
24.2. 2018
Lisäys:
Kuuntelin väitötilaisuuden 15.3. 2018 Mölndalin Yliopistollisen sairaalan uudessa valoisassa R-salissa nyt perjantai aamupäivällä. Väitös käytiin englanniksi, koska ulkomaalaisia osallistui tilaisuuteen. Suomalainen vastaväittelijäprofessori toimii Tukholman Karoliinisessa instituutissa onkologian alalla. Myös onkologian alan edustajia oli tutkintolautakunnassa, samoin psykiatrian alan edustaja, koska kyse oli neurodegeneratiivisten tautien diagnostiikan biomerkitsijöistä.
Ehdin viime viikolla katsoa teesikirjasta osan johdantoa. Johdannosta minua erityisesti kiinnostava osa koski ubikitiini-proteosomi asian puolta, joihin nämä TRIM-proteiinit joita katselen, myös kuuluvat. Suomennan siitä kappaleesta asiaa, nyt kun sain aamulla väitöskirjan käsiini.
(Sivulta 12) Kappaleesta 1.2.2. The Ubiquitin-Proteosome system
Ubikitiini-proteosomisysteemi on eräs evolutionaalinen konservoitunut aineenvaihdunnallinen tie, jossa proteiineja hajoitetaan siten, että kohdeproteiini saa ubikitiinileiman ("osoitelapun") ja siten kohdennetaan proteiinisilppuriin , 26S-proteosomiin, jossa se hajoaa - (Huom, kommenttini: ei siis olemattomaksi eikä ilmeisesti energiaksikaan, vaan esim aminohapoiksi, joita voi hyödyntää, sillä solulla on kova tarve näistä hyvistä alkutekijöistä esim tRNA hakulaitteella niitä viedään peptidisynteesiin ja kudosten uudistamiseen).
Tämä "postimerkki osoitelappu" ubikitiini eristettiin ensimmäisen kerran naudan kateenkorvasta.
Goldstein et al. 1975. Samoihin aikoihin eristettiin ubikitiinin polypeptidi
Schlesinger DH, Goldstein G, Niall HD. The complete aminoacid sequencde of ubiquitin, an adenylate cyclase stimulating polypeptide probably universal in living cells 1975.
(Muuten - katselin sen propeptidin rakenteen: siinä on 9 kertaa sama yhden ubikitiinin jakso toistuneena: siis "Ub-Ub-Ub-Ub-Ub-Ub-Ub-Ub-Ub").
Sitten havaittiin ATP:stä riippuva proteolyyttinen systeemi:
Herschko A, Ciechanover A et al. Resolution of the ATP-dependent proteolytic system from reticulocytes; a component that interacts with ATP (1979).
Wilkinson KD, Urban et al. Ubiquitin is the ATP-dependent proteolysis factor 1 of rabbit reticulocytes (1980).
(Siis Ubikitiini-proteosomi järjestelmä VAATII energiaa EIKÄ TUOTA energiaa, se siis siivoaa, hyödyntää ja siihen se tarvitsee palkan. Loogista.)
Järjestelmän silppuriosa 26S proteosomi on proteiinikompleksi, jossa on sylinterinomainen 20S ydinpartikkeli ja sillä on entsymaattinen kyky: se on proteaasi, joka tuottaa lyhyitä peptidipätkiä. Sitten siinä on porttiosa, 19S alayksikkö, johon kuuluu ubikitiinireseptori ja deubikitinaatioaktiivisuus, eli se irrottaa ubikitiinin, jota voi käyttää uudestaan.
(Otan linkin netistä:http://jme.endocrinology-journals.org/content/34/2/281/F1.expansion.html )

On olemassa kuitenkin 20S ja 26S proteosomi ubikitiinistä riippumatontakin proteiinin hajoamista.
Väitöskirja antaa viitteen:
(Takaisin Simon Sjödinin väitöskirjan johdannon tekstiin):
Ubikitiini (Ub) konjugoituu proteiinisubstraatteihin E1-aktivoivien entsyymien , E2-konjugoivien entsyymien ja E3-ligeeraavien entsyymien avustamana. Tällainen prosessi vaatii ATP-energiaa ja tuottaa , tioliesteri välituotteena, sillan ubikitiinin C-terminaalisen glysiinin ja substraatin lysiinin (K) väliin. E3 ensikädessä vastaa siitä, mihin substraattiin ubikitiini pitää kovalentisti liittää. Ihmisgenomi koodaa kahta E1-ubikitiiniä aktivoivaa entsyymiä ja noin 20 E2- konjugoivaa entsyymiä ja yli 600 E3 ligaasia. Lisäksi ubikitinaatio on käänteinen , reversibeli, ja deubikitinaaseilla voidaan Ub irrottaa substraateistaan.
Ubikitiini on posttranslationaalinen modifikaatio ja esiintyy monoubikitiininä ja polyubikitiininä. Ubikitiinillä on muitakin tehtäviä kuin vain toimia substraattien osoitelappuna proteiinisilppuriin
( proteosomiin). Se esim. säätelee endosytoosia ja solureseptorien hajoitusta ja säätelee myös transkriptiota.
---
( Kommentti. Arvelen näin: Koska propeptidi on eräänlainen resurssi, siitä saa koostettua erilaisia pätkiä, joilla on joka koolla on oma koodaava merkityksensä ja antimikrobiaalisessa funktiossa ne voivat vahvistaa merkkaavaa signaalia asettumalla rinnakkain tuhottavaan kohteeseen- toinen tie on että ne jakaantuvat heti 9 kappaleeksi ja sitten takaisin koostuvat eri pitoisuuksiin- sitä yleensä kuvatan kaavakuvissa, mutta toinenkin mahdollisuus on, vaatii tietysti spesifisen entyymin joka tekee esim 4 ub pituutta- tässä vain ajatus).
Jatkan taas eri TRIM- E3 ligaasien luettelon tekemistä
Päivän väitäkirjassa on monin paikoin lisää tästä ubikitiinipuolesta. Otan esiin myöhemmin.
Väitöskirjassa keskityttiin diagnostiikan tehostamiseen neurodegeneratiivisissa taudeissa ja parhaiden mahdollisten biomerkitsijöiden löytämiseen.
Omasta puolestani esitin ehdotuksen, että tähän ubikitiinipuoleen ja noihin 600 E3 ligaasiin kohdistettaisiin huomiota, sillä ne ovat aika pitkiä aminohappoketjuja ja vaativat ATP:tä toimiakseen ja nykyihminen on iän loppupuolella usein proteiinienergia-malnutritiossa- mainitsematta ollenkaan sitä stekytemutageeniproteiinin paljoutta jokaikäisten ravinnossa- mistä ei yksikään E3 ligaasi saa edes silppurin kauttakaan mitään hyödyllistä tRNA:lle esiin. Ravintosuosituksissa pitäisi hyvien alkuaineiden lisäksi mainita, miten niistä saa proteiinit imeytettyä kehoon tarpeeksi hyväkuntoisena, jotta keholle on edes vähän hyötyä ruoan proteiinista. Muutenhan kaikki vain muuttuu neutraalirasvaksi ja kuonatypeksi. Proteiiniapparaatti joka korjaa kehoa, voi kitua "oikean" proteiinin puutetta- ja samalla on vain suuri typpirasitus, typpiainesjätettä ja epäorgaansita fosfaattia metaboliseen rasitukseen asti. uusi tieto minulle oli se että neurodegeneratiivisissa kappaleissa ja aggrekaateissa on ylimääräistä ubikitiiiniäkin. Siksi olisi tuo alue proteostaasista mielestäni avainasemassa tutkimuskohteena,koska sen selvittämisestä voi jopa lähteä parantavia linjoja liikkelle luonnostaan ( innate immunity). Ubikitiini marköörinä on tietysti hyvin tärkeä.
pääpainotus oli diagnostiikaqssa ja biomarköörienlöytämisessä, eikävarinaisesti ubikitiiniproteosomijärjestelmässä, muta otan sen mitä löydän tästä jälkimmäisestä asiasta.
Fakultetsopponent Janne Lehtiö, professor KI STH.
http://hdl.handle.net/2077/54533
Suomennosta abstraktin sisällöstä:
Neurodegeneraatio on prosessi, jossa neuronisoluja katoaa ja oireista voidaan päätellä, miltä keskushermostoalueelta oireet johtuvat. Neurodegeneratiivisiin tauteihin lasketaan AD (Alzheimerin tauti) , PD (Parkinsonin tauti) ja FTD (frontotemporaalinen dementia) ja niissä kaikissa kertyy proteiiniaggrekaatteja, mikä viittaa näiden tuotannon tai vaihtumisen/hajottamisen olevan epäkunnossa. Täten toimimaton proteostaasi (proteiinien epätasapaino) näyttäisi olevan kyseessä neurodegeneratiivisissa taudeissa.
AD- taudissa on vähentynyt endosyyttisten ja autofagisten rakkuloiden vaihtuvuus ja niin kertyy aivokudokseen endolysosomaalisia valkuaisaineita ja käyttymätöntä ubikitiiniä.
PD- taudissa on viitettä lysosomien vikatoiminnasta tautiriskien ja geneettisten muuntumisten liittyessä lysosomaalisiin kertymishäiriöihin ja taudin vikuuttamien alueitten lysosomaalisten proteiinien vähentyneestä esiintymästä.
FTD:ssä tautia aiheuttavat mutaatiot ja geneettiset riskitekijät viittaavat autofagisen ja endosomaalisen järjestelmän muuntuneen toiminnan olevan osallisena patogeneesissä.
Väitöstyö selvittää proteostaasin vikatoimintaa aivoselkäydinnesteestä käsin- ja tähän kehitetään mittausmenetelmä. Siinä mitataan autofagisen ja endolysosomaalisen systeemin ja ubikitiini-proteosomi-systeemin proteiineja.
1) LAMP2, lysosomaali kalvoproteiini
2) kokopitkä ubikitiini ( joka on 9 ubikitiini-monomeerin pituinen)
3) endolysosomaalisten proteiinien paneli
Kehitetyllä menetelmällä analysoitiin mainittujen tautien (myös FTD alatyyppien ja FTD mutaation kantajien) likvornäytteet.
Johtopäätöksiä mm.
Mahdollisia biomerkitsijöinä voisi toimia likvorin ubikitiini AD-taudissa (korkea) ja endolysosomaaliset proteiinit ja ubikitiini PD-taudissa (matalat) kontrolleihin verrattuna. FTD taudissa löydöt dysfunktionaalisista proteiineista eivät olleet välittömästi havaittavissa.
24.2. 2018
Lisäys:
Kuuntelin väitötilaisuuden 15.3. 2018 Mölndalin Yliopistollisen sairaalan uudessa valoisassa R-salissa nyt perjantai aamupäivällä. Väitös käytiin englanniksi, koska ulkomaalaisia osallistui tilaisuuteen. Suomalainen vastaväittelijäprofessori toimii Tukholman Karoliinisessa instituutissa onkologian alalla. Myös onkologian alan edustajia oli tutkintolautakunnassa, samoin psykiatrian alan edustaja, koska kyse oli neurodegeneratiivisten tautien diagnostiikan biomerkitsijöistä.
Ehdin viime viikolla katsoa teesikirjasta osan johdantoa. Johdannosta minua erityisesti kiinnostava osa koski ubikitiini-proteosomi asian puolta, joihin nämä TRIM-proteiinit joita katselen, myös kuuluvat. Suomennan siitä kappaleesta asiaa, nyt kun sain aamulla väitöskirjan käsiini.
(Sivulta 12) Kappaleesta 1.2.2. The Ubiquitin-Proteosome system
Ubikitiini-proteosomisysteemi on eräs evolutionaalinen konservoitunut aineenvaihdunnallinen tie, jossa proteiineja hajoitetaan siten, että kohdeproteiini saa ubikitiinileiman ("osoitelapun") ja siten kohdennetaan proteiinisilppuriin , 26S-proteosomiin, jossa se hajoaa - (Huom, kommenttini: ei siis olemattomaksi eikä ilmeisesti energiaksikaan, vaan esim aminohapoiksi, joita voi hyödyntää, sillä solulla on kova tarve näistä hyvistä alkutekijöistä esim tRNA hakulaitteella niitä viedään peptidisynteesiin ja kudosten uudistamiseen).
Tämä "postimerkki osoitelappu" ubikitiini eristettiin ensimmäisen kerran naudan kateenkorvasta.
Goldstein et al. 1975. Samoihin aikoihin eristettiin ubikitiinin polypeptidi
Schlesinger DH, Goldstein G, Niall HD. The complete aminoacid sequencde of ubiquitin, an adenylate cyclase stimulating polypeptide probably universal in living cells 1975.
(Muuten - katselin sen propeptidin rakenteen: siinä on 9 kertaa sama yhden ubikitiinin jakso toistuneena: siis "Ub-Ub-Ub-Ub-Ub-Ub-Ub-Ub-Ub").
Sitten havaittiin ATP:stä riippuva proteolyyttinen systeemi:
Herschko A, Ciechanover A et al. Resolution of the ATP-dependent proteolytic system from reticulocytes; a component that interacts with ATP (1979).
Wilkinson KD, Urban et al. Ubiquitin is the ATP-dependent proteolysis factor 1 of rabbit reticulocytes (1980).
(Siis Ubikitiini-proteosomi järjestelmä VAATII energiaa EIKÄ TUOTA energiaa, se siis siivoaa, hyödyntää ja siihen se tarvitsee palkan. Loogista.)
Järjestelmän silppuriosa 26S proteosomi on proteiinikompleksi, jossa on sylinterinomainen 20S ydinpartikkeli ja sillä on entsymaattinen kyky: se on proteaasi, joka tuottaa lyhyitä peptidipätkiä. Sitten siinä on porttiosa, 19S alayksikkö, johon kuuluu ubikitiinireseptori ja deubikitinaatioaktiivisuus, eli se irrottaa ubikitiinin, jota voi käyttää uudestaan.
(Otan linkin netistä:http://jme.endocrinology-journals.org/content/34/2/281/F1.expansion.html )
On olemassa kuitenkin 20S ja 26S proteosomi ubikitiinistä riippumatontakin proteiinin hajoamista.
Väitöskirja antaa viitteen:
- Baugh JM, Viktorova EG. PROTEOSOMES CAN DEGRADE A SIGNIFICANT PROPORTION OF CELLULAR PROTEINS INDEPENDENT OF UBIQITINATION.
- ( Etsin tässä välillä tämän viitteen- Tätä pitää myöhemmin katsoa tarkemmin).
- Abstract, The critical role of the ubiquitin-26S proteasome system in regulation of protein homeostasis in eukaryotes is well established. In contrast, the impact of the ubiquitin-independent proteolytic activity of proteasomes is poorly understood.
- Through biochemical analysis of mammalian lysates, we find that the 20S proteasome, latent in peptide hydrolysis, specifically cleaves more than 20% of all cellular proteins. Thirty intrinsic proteasome substrates (IPSs) were identified and in vitro studies of their processing revealed that cleavage occurs at disordered regions, generating stable products encompassing structured domains. The mechanism of IPS recognition is remarkably well conserved in the eukaryotic kingdom, as mammalian and yeast 20S proteasomes exhibit the same target specificity.
- Further, 26S proteasomes specifically recognize and cleave IPSs at similar sites, independent of ubiquitination, suggesting that disordered regions likely constitute the universal structural signal for IPS proteolysis by proteasomes.
- Finally, we show that proteasomes contribute to physiological regulation of IPS levels in living cells and the inactivation of ubiquitin-activating enzyme E1 does not prevent IPS degradation.
- Collectively, these findings suggest a significant contribution of the ubiquitin-independent proteasome degradation pathway to the regulation of protein homeostasis in eukaryotes.
Ubikitiini (Ub) konjugoituu proteiinisubstraatteihin E1-aktivoivien entsyymien , E2-konjugoivien entsyymien ja E3-ligeeraavien entsyymien avustamana. Tällainen prosessi vaatii ATP-energiaa ja tuottaa , tioliesteri välituotteena, sillan ubikitiinin C-terminaalisen glysiinin ja substraatin lysiinin (K) väliin. E3 ensikädessä vastaa siitä, mihin substraattiin ubikitiini pitää kovalentisti liittää. Ihmisgenomi koodaa kahta E1-ubikitiiniä aktivoivaa entsyymiä ja noin 20 E2- konjugoivaa entsyymiä ja yli 600 E3 ligaasia. Lisäksi ubikitinaatio on käänteinen , reversibeli, ja deubikitinaaseilla voidaan Ub irrottaa substraateistaan.
Ubikitiini on posttranslationaalinen modifikaatio ja esiintyy monoubikitiininä ja polyubikitiininä. Ubikitiinillä on muitakin tehtäviä kuin vain toimia substraattien osoitelappuna proteiinisilppuriin
( proteosomiin). Se esim. säätelee endosytoosia ja solureseptorien hajoitusta ja säätelee myös transkriptiota.
---
( Kommentti. Arvelen näin: Koska propeptidi on eräänlainen resurssi, siitä saa koostettua erilaisia pätkiä, joilla on joka koolla on oma koodaava merkityksensä ja antimikrobiaalisessa funktiossa ne voivat vahvistaa merkkaavaa signaalia asettumalla rinnakkain tuhottavaan kohteeseen- toinen tie on että ne jakaantuvat heti 9 kappaleeksi ja sitten takaisin koostuvat eri pitoisuuksiin- sitä yleensä kuvatan kaavakuvissa, mutta toinenkin mahdollisuus on, vaatii tietysti spesifisen entyymin joka tekee esim 4 ub pituutta- tässä vain ajatus).
Jatkan taas eri TRIM- E3 ligaasien luettelon tekemistä
Päivän väitäkirjassa on monin paikoin lisää tästä ubikitiinipuolesta. Otan esiin myöhemmin.
Väitöskirjassa keskityttiin diagnostiikan tehostamiseen neurodegeneratiivisissa taudeissa ja parhaiden mahdollisten biomerkitsijöiden löytämiseen.
Omasta puolestani esitin ehdotuksen, että tähän ubikitiinipuoleen ja noihin 600 E3 ligaasiin kohdistettaisiin huomiota, sillä ne ovat aika pitkiä aminohappoketjuja ja vaativat ATP:tä toimiakseen ja nykyihminen on iän loppupuolella usein proteiinienergia-malnutritiossa- mainitsematta ollenkaan sitä stekytemutageeniproteiinin paljoutta jokaikäisten ravinnossa- mistä ei yksikään E3 ligaasi saa edes silppurin kauttakaan mitään hyödyllistä tRNA:lle esiin. Ravintosuosituksissa pitäisi hyvien alkuaineiden lisäksi mainita, miten niistä saa proteiinit imeytettyä kehoon tarpeeksi hyväkuntoisena, jotta keholle on edes vähän hyötyä ruoan proteiinista. Muutenhan kaikki vain muuttuu neutraalirasvaksi ja kuonatypeksi. Proteiiniapparaatti joka korjaa kehoa, voi kitua "oikean" proteiinin puutetta- ja samalla on vain suuri typpirasitus, typpiainesjätettä ja epäorgaansita fosfaattia metaboliseen rasitukseen asti. uusi tieto minulle oli se että neurodegeneratiivisissa kappaleissa ja aggrekaateissa on ylimääräistä ubikitiiiniäkin. Siksi olisi tuo alue proteostaasista mielestäni avainasemassa tutkimuskohteena,koska sen selvittämisestä voi jopa lähteä parantavia linjoja liikkelle luonnostaan ( innate immunity). Ubikitiini marköörinä on tietysti hyvin tärkeä.
lördag 17 februari 2018
Ihmisen koodin kirjoituskone. Onko keholla "kirjoituskoneallergiaa?"
DOI
https://www.biorxiv.org/content/early/2018/01/05/243345
(PS. Tulee mieleen analogisesti ubikitiini, onkohan se jokin vanha eliö, olio, bakteeri, kun sillä on propeptidi 9 toistosekvenssiä 9x76, ja sellainen syöjäproteosomihahmo kuuluu sen järjestelmään jolla on kuin pää ja häntä, mutta olion hienontamat tuotteet hyödynnetään kehomiljöössä
Ubikitiinin esiproteiinista tulee 9 kpl monoubikitiiniä) Ihmiskeho on ottanut sen hyötykäyttöön ja se on hyvin monipuolinen värkki: Niitä osasia voi asettaa peräkkäin tai vierekkäin tilanteen mukaan vahventaen antivirusvastetta).
This was all puzzling until scientists realized the unique sequences in between the repeats matched the DNA of viruses—specifically viruses that prey on bacteria. It turns out CRISPR is one part of the bacteria’s immune system, which keeps bits of dangerous viruses around so it can recognize and defend against those viruses next time they attack. The second part of the defense mechanism is a set of enzymes called Cas (CRISPR-associated proteins), which can precisely snip DNA and slice the hell out of invading viruses. Conveniently, the genes that encode for Cas are always sitting somewhere near the CRISPR sequences.
https://www.nature.com/articles/srep37895
The
international race to bring human genome editing into widespread use in
clinical medicine is moving fast. On Jan 23, the National Institutes of
Health Common Fund launched its Somatic Cell Genome Editing programme,
committing approximately US$190 million of funding over the next 6
years to propel development of genome editing into medical practice. A
worthy effort, but the USA and Europe still trail behind China. As many
as 86 patients in China have already had their genes altered as part of
clinical trials to treat a range of diseases, including solid cancers.
Findings from these studies are yet to be reported in peer-reviewed
journals. Worryingly, serious criticisms have been levelled at China for forging ahead with these trials without sufficient regulatory review.
Investigators
of first in-human genome editing clinical trials in the USA and Europe
have applied to the US Food and Drug Administration and the European
Medicines Agency, respectively, to begin enrolling patients this year.
Originally planned to start in 2016, these studies have, in part, been
hampered by regulatory and safety concerns. Concerns that
are by no means unwarranted—as the technology advances, questions about
safety will undoubtedly be raised. A study
published on the bioRxiv preprint server this year showed pre-existing
adaptive immunity against bacterial-derived Cas9 in human serum, which
could result in patients having immune reactions against the
gene-editing machinery. Additionally, although all trials so far
registered modify genes ex vivo in cells that are then reintroduced into
patients, the next logical step is directly editing genes in vivo. A 2017 report
of an animal study using an in vivo CRISPR/Cas9 system showed an
unexpected number of off-target mutations, an important signal that
further research is needed before in vivo gene editing techniques can be
introduced into humans.Human genome editing is no
longer a concept confined to the pages of futuristic science fiction
novels—modifying genetic code is here now and is advancing rapidly.
Globally, regulators and investigators must work together to ensure
oversight of the development of gene editing technologies. Regulations
must not only keep up and anticipate future applications, but also
facilitate swift and safe implementation of the technology in the clinic. https://www.biorxiv.org/content/early/2018/01/05/243345
(PS. Tulee mieleen analogisesti ubikitiini, onkohan se jokin vanha eliö, olio, bakteeri, kun sillä on propeptidi 9 toistosekvenssiä 9x76, ja sellainen syöjäproteosomihahmo kuuluu sen järjestelmään jolla on kuin pää ja häntä, mutta olion hienontamat tuotteet hyödynnetään kehomiljöössä
Ubikitiinin esiproteiinista tulee 9 kpl monoubikitiiniä) Ihmiskeho on ottanut sen hyötykäyttöön ja se on hyvin monipuolinen värkki: Niitä osasia voi asettaa peräkkäin tai vierekkäin tilanteen mukaan vahventaen antivirusvastetta).
CRISPR/Cas9 comes from strep bacteria...
CRISPR is actually a naturally-occurring, ancient defense mechanism found in a wide range of bacteria. As far as back the 1980s, scientists observed a strange pattern in some bacterial genomes. One DNA sequence would be repeated over and over again, with unique sequences in between the repeats. They called this odd configuration “clustered regularly interspaced short palindromic repeats,” or CRISPR.This was all puzzling until scientists realized the unique sequences in between the repeats matched the DNA of viruses—specifically viruses that prey on bacteria. It turns out CRISPR is one part of the bacteria’s immune system, which keeps bits of dangerous viruses around so it can recognize and defend against those viruses next time they attack. The second part of the defense mechanism is a set of enzymes called Cas (CRISPR-associated proteins), which can precisely snip DNA and slice the hell out of invading viruses. Conveniently, the genes that encode for Cas are always sitting somewhere near the CRISPR sequences.
https://www.nature.com/articles/srep37895
Article
|
Open
A CRISPR-Cas9 Assisted Non-Homologous End-Joining Strategy for One-step Engineering of Bacterial Genome
- Scientific Reports volume 6, Article number: 37895 (2016)
- doi:10.1038/srep37895
fredag 16 februari 2018
TRIMproteiiniperhe on jaettavissa funktionaalisiin alaryhmiin domeenikoostumuksen mukaan
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5314404/
Activated
macrophages play an important role in many inflammatory diseases
including septic shock and atherosclerosis. However, the molecular
mechanisms limiting macrophage activation are not completely understood.
Members of the tripartite motif (TRIM) family have recently emerged as important players in innate immunity and antivirus. Here, we systematically analyzed mRNA expressions of representative TRIM molecules in human THP1-derived macrophages activated by different Toll-like receptor (TLR) ligands.
Twenty-nine TRIM members were highly induced by one or more TLR ligands, among which 19 of them belong to TRIM C-IV subgroup.
Besides TRIM21, TRIM22 and TRIM38 were shown to be upregulated by TLR3 and TLR4 ligands as previous reported, we identified a novel group of TRIM genes (TRIM14, 15, 31, 34, 43, 48, 49, 51 and 61) that were significantly up-regulated by TLR3 and TLR4 ligands.
In contrast, the expression of TRIM59 was down-regulated by TLR3 and TLR4 ligands in both human and mouse macrophages.
The alternations of the TRIM proteins were confirmed by Western blot. Finally, overexpression of TRIM59 significantly suppressed LPS-induced macrophage activation, whereas siRNA-mediated knockdown of TRIM59 enhanced LPS-induced macrophage activation.
Taken together, the study provided an insight into the TLR ligands-induced expressions of TRIM family in macrophages.
Members of the tripartite motif (TRIM) family have recently emerged as important players in innate immunity and antivirus. Here, we systematically analyzed mRNA expressions of representative TRIM molecules in human THP1-derived macrophages activated by different Toll-like receptor (TLR) ligands.
Twenty-nine TRIM members were highly induced by one or more TLR ligands, among which 19 of them belong to TRIM C-IV subgroup.
Besides TRIM21, TRIM22 and TRIM38 were shown to be upregulated by TLR3 and TLR4 ligands as previous reported, we identified a novel group of TRIM genes (TRIM14, 15, 31, 34, 43, 48, 49, 51 and 61) that were significantly up-regulated by TLR3 and TLR4 ligands.
In contrast, the expression of TRIM59 was down-regulated by TLR3 and TLR4 ligands in both human and mouse macrophages.
The alternations of the TRIM proteins were confirmed by Western blot. Finally, overexpression of TRIM59 significantly suppressed LPS-induced macrophage activation, whereas siRNA-mediated knockdown of TRIM59 enhanced LPS-induced macrophage activation.
Taken together, the study provided an insight into the TLR ligands-induced expressions of TRIM family in macrophages.
Macrophages
are the major components of innate immunity that enable the body to
combat bacteria and other pathogens.
However, over-activation of macrophages plays a central role in a variety of inflammatory diseases, such as septic shock, atherosclerosis, arthritis and inflammatory bowel diseases. In these disease settings, activated macrophages elaborate a large array of cytokines, growth factors and proteolytic enzymes that are critical for tissue damage and repair1,2.
Macrophages are activated in response to the pathogen-associated molecular patterns by various pattern-recognition receptors (PRRs), such as the Toll-like receptors (TLRs) and the RIG-I-like receptors (RLR)3,4.
There are 13 TLRs that sense various pathogen components and trigger intracellular signaling pathways that eventually mediate the induction of inflammatory cytokines, chemokines and type I interferons, which are critical for antimicrobial activity4,5. The molecular mechanisms of regulation of macrophage activation in response to TLR ligands have been largely unknown.
However, over-activation of macrophages plays a central role in a variety of inflammatory diseases, such as septic shock, atherosclerosis, arthritis and inflammatory bowel diseases. In these disease settings, activated macrophages elaborate a large array of cytokines, growth factors and proteolytic enzymes that are critical for tissue damage and repair1,2.
Macrophages are activated in response to the pathogen-associated molecular patterns by various pattern-recognition receptors (PRRs), such as the Toll-like receptors (TLRs) and the RIG-I-like receptors (RLR)3,4.
There are 13 TLRs that sense various pathogen components and trigger intracellular signaling pathways that eventually mediate the induction of inflammatory cytokines, chemokines and type I interferons, which are critical for antimicrobial activity4,5. The molecular mechanisms of regulation of macrophage activation in response to TLR ligands have been largely unknown.
Tripartite
motif (TRIM) proteins contain a RING finger, one or two B-box motifs
and a coiled-coil motif, and are involved in many biological processes
including innate immunity, viral infection, carcinogenesis and
development6.
There are over 70 members of TRIM protein family described in humans7. Recently, several systematic analyses suggest that many TRIM proteins are implicated in the regulation of innate immune pathways and anti-viral activities8,9,10,11. For example, Carthagena et al. identified 27 of the 72 human TRIM genes are sensitive to interferon (IFN) by performing a systematic analysis of TRIM gene expressions in human primary lymphocytes and monocyte-derived macrophages in response to IFNs10.
In addition, Rajsbaum et al. found that the genes encoding a subset of TRIM proteins located on chromosome 7 were up-regulated by type I IFN in macrophages/DC, suggesting that they may have anti-viral functions11.
TRIM8 negatively regulates PIAS3-mediated repression of NF-κB by inducing translocation of PIAS3 from nucleus to cytoplasm as well as its turnover12,13,14, whereas TRIM16 (also known as EBBP) was reported to promote IL-1β secretion. TRIM22 is involved in anti-viral pathways by activating NF-κB signaling15,16,17,18. TRIM30 induces the lysosomal degradation of TAB2 and TAB3, thereby negatively regulating NF-κB induction in the LPS-triggered TLR4 signaling pathway19. TRIM21 negatively regulates TLR3, −4, −7, and −9 and RLR signaling pathways by modulating the activities of IKKs and interferon regulatory factors (IRFs)20,21. TRIM27 targets all IKKs and negatively regulates the PRR pathways21,22. CARD domain ubiquitination by TRIM25 is essential for RIG-I-mediated type I interferon induction21,23. TRIM56 facilitates double-strand DNA-stimulated interferon induction by ubiquitination of STING (stimulator of interferon genes)21,24. However, the functions of most of TRIM family members remain to be characterized.
There are over 70 members of TRIM protein family described in humans7. Recently, several systematic analyses suggest that many TRIM proteins are implicated in the regulation of innate immune pathways and anti-viral activities8,9,10,11. For example, Carthagena et al. identified 27 of the 72 human TRIM genes are sensitive to interferon (IFN) by performing a systematic analysis of TRIM gene expressions in human primary lymphocytes and monocyte-derived macrophages in response to IFNs10.
In addition, Rajsbaum et al. found that the genes encoding a subset of TRIM proteins located on chromosome 7 were up-regulated by type I IFN in macrophages/DC, suggesting that they may have anti-viral functions11.
TRIM8 negatively regulates PIAS3-mediated repression of NF-κB by inducing translocation of PIAS3 from nucleus to cytoplasm as well as its turnover12,13,14, whereas TRIM16 (also known as EBBP) was reported to promote IL-1β secretion. TRIM22 is involved in anti-viral pathways by activating NF-κB signaling15,16,17,18. TRIM30 induces the lysosomal degradation of TAB2 and TAB3, thereby negatively regulating NF-κB induction in the LPS-triggered TLR4 signaling pathway19. TRIM21 negatively regulates TLR3, −4, −7, and −9 and RLR signaling pathways by modulating the activities of IKKs and interferon regulatory factors (IRFs)20,21. TRIM27 targets all IKKs and negatively regulates the PRR pathways21,22. CARD domain ubiquitination by TRIM25 is essential for RIG-I-mediated type I interferon induction21,23. TRIM56 facilitates double-strand DNA-stimulated interferon induction by ubiquitination of STING (stimulator of interferon genes)21,24. However, the functions of most of TRIM family members remain to be characterized.
In
the present study, we systematically profiled the expressions of TRIM
gene family in human THP1-derived macrophages activated by different TLR
ligands. The up-regulated or down-regulated TRIM genes were further
confirmed by quantitative real-time polymerase chain reaction (qRT-PCR)
and Western blot analysis. The function of TRIM59 in macrophage
activation was further studied.
Tuore väitöskirja syövän genomisista muutoksista (2018, A-M BABAK)
Thesis
CANCER februari 2018
http://hdl.handle.net/2077/54530
| Titel: | Comprehensive analysis of structural genomic alterations in cancer |
| Övriga titlar: | Computational approaches for identifying cancer driver events |
| Författare: | Alaei-Mahabadi, Babak |
| Publikationstyp: | Doctoral thesis |
| Nyckelord: | Somatic structural variations viral integrations gene fusions somatic genomic alterations |
978-91-629-0423-4
(PDF)
978-91-629-0422-7 (PRINT)
978-91-629-0422-7 (PRINT)
Suomennosta
abstraktin sisällöstä:
Normaalin solun
transformoituminen syöpäsoluksi käsittää sellaisten somaattisten
DNA-muutosten kertymistä, joista tulee syövän kasvua ja
elossapysymisetuja. Tällaiset genomsiet muutokset ovat erilaisia
mallinsa ja laajuutensa suhteen, niitä on yksittäisen nukleotidin
variantteja (SNV) pieniä insertioita ( genomisen palasen
liittymiä) tai deleetioita (genomisen palasen poistumaa)
(indels) tai vieraan DNA-materiaalin settumaan genomiin, esim
viruksen DNA:ta.
-
The transformation of a normal cell into a cancer cell involves the accumulation of somatic DNA alterations that confer growth and survival advantages. These genomic alterations can be different in terms of pattern and size, comprising single nucleotide variants (SNVs), small insertions or deletions (indels), structural variations (SVs) or foreign DNA insertions such as viral DNA.
Syöpägenomille on
tyypillistä lukuisat sellaiset muutokset, joista vain vähäinen
murto-osa on tuumorin evoluutiossa positiivisesti valiutuneita
tapahumien käynnistäjiä. Hyvin perusteelliset laajat ja nopeat
genomiset sekvenssoinnit ovat mahdollistaneet somaattisten
DNA-muutosten systemaattisen kartoituksen tuhansista
tuumorigenomeista. Tämän tyyppisestä tietoaineistosta on
perusteellisesti tutkittu mutaatioita ja on löydetty monia uusia
geenejä, jotka osallisuvat kasvaimen kehittymiseen. Mutta
tietämys yksittäisten nukleotidivarianttien osuudesta syövässä
on edelleen rajallista.
-
Cancer genomes typically harbor numerous such changes, of which only small fractions are driver events that are positively selected for during the evolution of the tumor. High throughput sequencing has enabled systematic mapping of somatic DNA alterations across thousands of tumor genomes. Mutations in particular have been thoroughly explored in this type of data, and this has implicated many new genes in tumor development. However, our knowledge remains more limited when it comes to the contribution of SVs to cancer.
Tässä
väitöskirjatyössä tutkijaryhmä käytti yleisesti saatavilla
olevaa geenitietämystä syövistä hankkiakseen lisää
ymmärtämystä tuumorin kehittymisestä.
-
In the present thesis, we made use of publicly available cancer genomics data to gain further insight into the role of structural genomic alterations in tumor development.
Virukset aiheuttavat
10-15 % kaikista ihmisen syövistä monien mekanismien välityksellä
ja yksi niistä on sellainen, missä rakenteelliset genomiset
muutokset johtuvat viruksen DNA:n integroitumisesta ( asettumisesta
geenijoukkoon) ihmisen genomiin. Niinpä ensimmäisessä
tutkimuksessa seulottiin virusgenomien integroitumisia
syöpägenomeihin.
-
Viruses cause 10-15% of all human cancers through multiple mechanisms, one of which is structural genomic changes due to viral DNA being integrated into the human genome.
Virusintegraation
havaitsemiseksi käytettiin RNA-sekvenssitietueita noin 4500
kasvaimesta, jotka edustivat 19 erilaista syöpätyyppiä. Tutkijat
havaitsivat tyypillisten toistuvien tapahtumin assosioitumista
tunnettuihin syöpägeeneihin ja niihin liittyi muuntunut geenin
ilmenemä. Yksinkertaiset rakenteelliset vaihtelut (SV) voivat
johtaa jonkin spesifisen syöpää aiheuttavan geenin kopioitten
lukumäärän moninkertaistumiseen kuten myös onkogeenien
fuusiomuodostumaan, mutta sen tyyppisten tapahtumien merkitystä
syövässä ei ole selvitetty tarpeeksi.
-
Thus, in the first study, we performed an unbiased screen for viral genomic integrations into cancer genomes. We developed a computational pipeline using RNA-Seq data from ~4500 tumors across 19 different cancer types to detect viral integrations. We found that recurrent events typically involved known cancer genes, and were associated with altered gene expression. SVs can lead to copy number amplification of specific cancer driver genes, as well as the formation of fusion oncogenes, but their importance in cancer beyond these types of events is underexplored.
Tutkijaryhmä
kartoitti rakenteellisia variaatioita ihmisen genomista käyttämällä
600 tuumorin kokogenomin sekvensoimista- näissä oli 10 eri
syöpätyyppiä – ja he selvittivät genomisen suhteen
rakenteellisten variaatioiden (SV) ja mRNA muutosten kesken. He
havaitsivat, että sellaiset tapahtumat antavat osansa ihmisen
tuumorien geeniexpressioon, mutta mittään uutta toistuvaa tuumoria
aiheuttavaa tapahtumaa he eivät löytäneet.
-
We mapped SVs to the human genome using whole genome sequencing data from 600 tumors across 18 different cancer types and investigated the global relationship between SVs and mRNA changes.
-
We found that such events often contribute to altered gene expression in human tumors, but we were not able to detect novel recurrent driver events.
Kohortin kokoa
lisättiin... ( 32 syöpälajia, noin 10 000 tuumoria) tarkoituksella
tunnistaa toistuvia rakenteellisen variaation aiheutamia
tapahtumia tuumoreista. Erityisesti tutkittiin niitä rakenteelisia
variaatioita, joiden odotettiin johtavan
promoottoirsubstituutiotapahtumiin, mikä on eräs tunnettu
mekanismi, jolla geeni aktivoituu syövässä. He löysivät useita
toistuvasti aktivoivia tapahtumia, joilla on mahdollisuus toimia
syövän muodostuksessa ”draiverina”.
Sitaatti 16.2. 2018
. Suomennosta abstraktista.
-
To increase the cohort size, we used a larger but lower resolution and more limited dataset, comprising of microarray based DNA copy number profiles from ~10,000 tumors across 32 cancer types, with the aim of identifying recurrent SV driver events in tumors. Specifically, we investigated SVs predicted to result in promoter substitution events, a known mechanism for gene activation in cancer, and found several recurrent activating events with potential cancer driver roles.
HPV-virusta
koskevissa tutkimusta saatiin huomionarvoinen löytö. Imisen
papilloomavirus integroituu geeneihin RAD51B ja ERBB2 ja genifuusiio
koskee geenjä NFF21.2, TIAM2 ja SCARB1 ja ne kaikki tunnetaan
syöpägeeneinä.
-
Notable among our findings in all the studies were human papillomavirus integrations in RAD51B and ERBB2 and gene fusions involving NFE2L2, TIAM2 and SCARB1, all being known cancer genes.
Yhteenvetona:
Runsaasta genomi-
ja transkriptomisekvenssianeistosta saadusta daatasta tutkijaryhmä
pystyi selkeästi ja laaja-alaisesti kartoittamaan
virusintegraatiota ja rakenteellista variaatiota syövässä.
Tiedon perusteella tunnistettiin useita geenejä, joilla on
mahdollista osuutta kasvaimen kehityksessä.
-
Taken together, massive amounts of genomic and transcriptomic sequencing data allowed us to comprehensively map viral integrations and structural variations in cancer, which led to the identification of several genes with potential roles in tumor development
Kts. väitöspäivän teksti: 2.3. 2018
torsdag 15 februari 2018
Luonnollinen immuniteetti. NFkB tumatekijästä . Interferonijärjestelmästä
Suomennosta artikkelista
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3945521/#
johdanto-osasta
Nisäkkäillä ja myös ihmisellä immuunivaste
jaetaan luonnolliseen ja adaptiiviseen mekanismiin. Luonnollinen
immuniteetti on ensimmäisenä ja jo valmis imuunipuolustus kehoon tulevia
patogeenejä vastaan ja se onkin kriittinen infektion kontrolloija
siinä ohikiitävässä ajanhetkessä, kun taas adaptiivisen
tehokkaan , spesifisen ja tarkan immuunivasteen pystyttämiseen menee oma
aikansa. ( Voisi verrata esim valtion automaattiseen
ohjuspuolustukseen, joka laukeaa heti, kun vihollinen lähestyy -
ja varsinaisen puolustuarmeijan kokoonkutsuminen ja tilanteen
analysointi vie olevaisia aikoja. Tästä jo huomaa miten tärkeä
on luonnollinen immuniteetti. Se heijastaa kehon kuntoa ja
resurssitilaa, valmiutta ennen kun infektio on ilmentynyt. ) .
-
In mammals the immune response is comprised of both innate and adaptive mechanisms. The innate response is the first line of defense against incoming pathogens and is crucial for controlling infection in the time it takes to mount an effective adaptive response 1.
Kriittiset luonnollisen immuniteetin vasteet viruksia
kohtaan käsittävät kehorakenteessa ilmeneviä proteiineja,
joilla on sisäsyntyisiä antimikrobi-ominaisuuksia ja kyky
kehkeyttää 1-tyypin interferonijärjestelmä (IFN-1), joka taas herättää uinuvia antivirusgeenejä.
Kehossa on PR-
reseptoreita, mallintunnistaja- reseptoreita. . Ne pystyvät
havaitsemaan, mitä laatua kehoon tullut mikrobi on tunnistamalla
patogeeniin liittyneen molekulaarisen mallin (PAMP = Pathogen
Associated Molecular Pattern).
Näitä reseptoreita ovat esim
Tollin reseptorin kaltaiset
reseptorit (TLR)
Retiinihapon indusoiman
geenin (RIG-1) kaltainen reseptori (RLR),
NOD:n kaltaiset reseptorit
(NLR) ( NOD on lyhennys sanoista: Nucleotide binding Oligomerization
Domain),
C-tyypin lektiinin kaltaiset
reseptorit (CLR).
-
The inducible antiviral response is initiated when pathogen associated molecular patterns (PAMPs) are recognized by pattern recognition receptors (PRRs), including Toll-like receptors (TLRs), retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) NOD-like receptors (NLRs) and C-type lectin receptors (CLRs) 5.
Näiden PR-reseptoreiden välittämästä herätteestä
liipaistuu esiin alavirran signalointiteitä erilaisten sopeuttavien
adaptoreina toimivien proteiinien kautta kohti tärkeää
kappaB-estäjäproteiinia (IkB) ja sitä fosforyloivaa kinaasia IKK
, jotka kuuluvat ns. NFkB-järjestelmään (Lue: n-f- kappa-b). NF
tarkoittaa tumafaktoria, jolla on oma säätelyjärjestelmänsä.
Tieto patogeeneista välitetään tälle järjestelmälle, joka
pystyy lähettämään edustajansa tumaan tietoa antamaan , että on virus kyseessä ja tarvitaan interferonia ja signaalinlähetys muills soluille , solun ulkopuolelle. IKK- ja
IkB- sanassa kirjain I tarkoittaa inhibiittoria, estäjää. IKK
tarkoittaa IkB-kinaasin estäjää. (Kinaasi on fosforyloiva
entsyymi).
Klassiset IKK entsyymit ( joissa on alayksikät alfa ja
beeta) vastaavat tumatekijän kappaB (kB) inaktivoitumsesta ja
siitä seurauksena tapahtuvasta proinflammatoristen eli
tulehduksellisten sytokiinien synteesistä.
Toiset kinaasit ( IKK:n
kaltaiset) fosforyloivat IRF3- proteiinin ( se on interferonia
säätelevä tekijä 3, joka pystyy DNA- transkriptioon tuman
puolella) . Myös IRF7 saa fosforylaation. Ne pystyvät
käskemään kirjoittamaan DNA:sta sanomaa, joka sitten tuottaa IFN-1-tyypin
interferoneja soluplasmaan ja solusta ulos.
Solun ulkopinnalla syntetisoitunut interferoni pystyy ulkokautta antamaan uuden signaalin soluun: Sille on
reseptori IFNR solupinnalla. Reseptori taas omaa soluliman puolella
päädyn ja pystyy heräytämään proteiinivaikuttajaketjun (JAK1,
TYK2 ovat kinaaseja ja fosforyloivat eli aktivoivat edelleen STAT1
ja STAT2, jotka ovat odotelleet inaktiiveina valmiina
solulimassa. Ne tekevät asiaankuuluvan komponentin IRF9 kanssa
kompleksin ( ISGF3) ja siirtyvät tumaan, jossa DNA on.
( IRF9
tarkoittaa "Interferonia säätelevä tekijä 9", mutta isompi kompleksi
ISGF3 on nimeltään
" interferonin stimuloima geenifaktori
3." Suorastaan tarkoittaa: "Tosi kysymyksessä" Nyt se on valmis menemään DNA:sta sijaitsevalle tehtäväpaikalleen
tumaan. Kromatiinissa on sille spesifisesti vastaavia kohtia ISRE,
" interferonista stimuloituva vaste-elementti".
Nyt kun
alkaa kirjoittautua genomikirjastosta interferonin indusoimia geenejä, niin
niitä on iso kirjo. Ja ne pystyvät puuttumaan viruksen
repliakaatio sykliin ihmistä puolustaen.
(Tässä on huomattava
että virukset ovat kehittäneet erilaisia menetelmiä välttää
(evaasio) interferonijärjestelmän aktivoituminen ja näiden erittäin monen
antivirusgeenin herääminen. Miltei jokainen virus keksii jonkin
evaasiokeinon, mutta yksi katalammista on iskeä niin, että solu
ei saa annettua interferonisignaaliaan solunsisätapahtumista
ulkopuolelle interferonin avulla..
-
The classical IKKs (IKKα/β) are responsible for the activation of nuclear factor kappa B (NF-κB) and subsequent pro-inflammatory cytokine synthesis [8] and [9], whereas the IKK-related kinases (TBK1/IKKε) phosphorylate the transcription factors IFN-regulatory factor (IRF) 3 and IRF7 required for IFN-I production [10] and [11]. Upon binding of IFN-I to its receptor, activation of JAK1 and TYK2 kinases results in phosphorylation of the transcription factors STAT1 and STAT2 to form a complex with IRF-9 known as IFN- ne stimulated gene factor 3 (ISGF3) [4] and [12]. This complex translocates to the nucleus and binds IFN-stimulated response elements (ISREs), ultimately resulting in the expression of a large set of IFN inducible genes (ISGs) which can directly interfere with the viral replication cycle [13] and [14].
onsdag 14 februari 2018
TRIM4, RNF87, kr. 7q22.1, Oxidatiivinen stressi ja mitokondria.
TRIM4
Mitä tiedetään TRIM-4
proteiinista? Geeni TRIM4 eli RNF87 sijaitsee kromosomissa t, q22.1. Porteiini lokalisoituu sytoplasmisesti. Funktio ei oltu vielä 2010 määritelty. Antiviraalia funktiota on osoitettu. E3 ubikitinaatiolla on myös solusignalointiin vaikuttavaa merkitystä. TRIM4 näyttää osallistuvan oxidatiivisen stressin vaikuttamaan apoptoosiin.
Suomennosta alla olevasta artikkelista
"On lisääntyvää näyttöä
siitä, että kohdeproteiinien modifioituminen ubikitiinillä ei
ainoastaan toimi turnover -koneistona
ubikitiini-proteiinisilppurijärjestelmässä (UPS) , vaan se on myös
kriittinen säätelijä erilaisissa signaaliteissä. Ubikitinaation
aikana E3ligaasi tunnistaa kohdeproteiinin ja päättää
ubikitiiniketjujen topologiasta. Tässä artikkelin työssä
tutkijat selvittivät TRIM4-proteiinin osuuttaa vetyperoksidin (H2O2)
indusoiman solukuoleman säätelyssä.
TRIM4 esiintyy eri tavalla
kehon eri kudoksissa ( kuten eri TRIM-proteiinit tapaavatkin tehdä.
Jokaisella nisitä on tyypillinen kirjonsa) , ja TRIM4 esiintyy
useimmissa syöpäkudoslinjoissa . Kun on selvitetty sen
subsellulaarista sijoittautumista , on osoittautunut, että TRIM4
muodostaa selvästi erillisen sytoplasmisen täplikkään
struktuurin, joka ohimenevästi tekee vuorovaikutusta
mitokondriaan. TRIM4:n ilmenemä aiheuttaa mitokondrioitten
aggrekoitumisen ja mitokondriaalisten vapaitten radikaalien
pitoisuuden nousua, jos vetyperoksidia on läsnä.
TRIM4 herkistää
solut vetyperoksidin aiheuttamalle solukuolemalle, apoptoosille, kun taas
TRIM4-poistogeenisyys oli vaikutukseltaan palauttava. TRIM4
vahvistaa mitokondrian kalvojenvälisen (tärkeän) potentiaalin
katoamista ja mitokondrian "hengen", sytokromi c:n
irtoamista mitokondriasta vetyperoksidin H2O2 vaikutuksesta.
Analysoitaessa TRIM4-
proteiinin kanssa vuorovaikutuksessa olevia proteiineja todetaan
peroxiredoxiini1 (PRX1) ja muita mitokondriaaliseen ja REDOX-
tasapainon säätelyyn osallistuvia proteiinia niiden joukossa.
TRIM4-proteiinin interaktio peroxiredoxiini-1:n kanssa on
kriittinen vetyperoksiidin indusoimassa solukuolemassa.
Kaiken kaikkiaan tämä
tutkimus viittaa TRIM4-proteiinilla olevan osaa oxidatiivisen
stressin indusoimassa solukuolemassa.
Free
Radic Biol Med. 2015 Dec;89:1036-48. doi:
10.1016/j.freeradbiomed.2015.10.425. Epub 2015 Oct 31.
TRIM4; a novel mitochondrial interacting RING E3
ligase, sensitizes the cells to hydrogen peroxide (H2O2) induced cell
death.
Tomar
D1, Prajapati
P2, Lavie
J3, Singh
K2, Lakshmi
S2, Bhatelia
K2, Roy
M2, Singh
R4, Bénard
G5, Singh
R6.
The emerging evidences suggest that posttranslational
modification of target protein by ubiquitin (Ub) not only regulate
its turnover through ubiquitin proteasome system (UPS) but is a
critical regulator of various signaling pathways. During
ubiquitination, E3 ligase recognizes the target protein and
determines the topology of ubiquitin chains. In current study, we
studied the role of TRIM4, a member of the TRIM/RBCC protein family
of RING E3 ligase, in regulation of hydrogen peroxide (H2O2) induced
cell death.
TRIM4 is expressed differentially in human tissues and
expressed in most of the analyzed human cancer cell lines. The
subcellular localization studies showed that TRIM4 forms distinct
cytoplasmic speckle like structures which transiently interacts with
mitochondria. The expression of TRIM4 induces mitochondrial
aggregation and increased level of mitochondrial ROS in the presence
of H2O2. It sensitizes the cells to H2O2 induced death whereas
knockdown reversed the effect. TRIM4 potentiates the loss of
mitochondrial transmembrane potential and cytochrome c release in the
presence of H2O2.
The analysis of TRIM4 interacting proteins showed its
interaction with peroxiredoxin 1 (PRX1), including other proteins
involved in regulation of mitochondrial and redox homeostasis. TRIM4
interaction with PRX1 is critical for the regulation of H2O2 induced
cell death. Collectively, the evidences in the current study suggest
the role of TRIM4 in regulation of oxidative stress induced cell
death.
KEYWORDS:
Cell death; H(2)O(2); Mitochondria; PRX1; TRIM4;
Ubiquitin E3 ligase
Muistiin suomennos 14.2.
2018
TRIM4 geenitieto:
PubMed
haku klo 22:51
- Official Symbol
- TRIM4provided by HGNC
- Official Full Name
- tripartite motif containing 4provided by HGNC
- Gene type
- protein coding
-
- Organism
- Homo sapiens
- Also known as
- RNF87
- Summary
- The protein encoded by this gene is a member of the tripartite motif (TRIM) family. The TRIM motif includes three zinc-binding domains, a RING, a B-box type 1 and a B-box type 2, and a coiled-coil region. The protein localizes to cytoplasmic bodies. Its function has not been identified. Alternatively spliced transcript variants that encode different isoforms have been described.[provided by RefSeq, Jul 2010]
- Expression
- Ubiquitous expression in ovary (RPKM 16.5), thyroid (RPKM 12.9) and 25 other tissues See more
- Orthologs
- all
Related articles(3) in PubMed
-
TRIM4; a novel mitochondrial interacting RING E3 ligase, sensitizes the cells to hydrogen peroxide (H2O2) induced cell death. Tomar D, et al. Free Radic Biol Med, 2015 Dec. PMID 26524401
(Ylläoleva artikkli suomeksi. . Vuodelta 2014)
MITEN TRIM4 on antiviraali?
Tämäkin artikkeli on vuodelta 2014.
-
TRIM4 modulates type I interferon induction and cellular antiviral response by targeting RIG-I for K63-linked ubiquitination. Yan J, et al. J Mol Cell Biol, 2014 Apr. PMID 24755855RIG-I is a pivotal cytoplasmic sensor that recognizes different species of viral RNAs. This recognition leads to activation of the transcription factors NF-κB and IRF3, which collaborate to induce type I interferons (IFNs) and innate antiviral response. In this study, we identified the TRIM family protein TRIM4 as a positive regulator of RIG-I-mediated IFN induction.Overexpression of TRIM4 potentiated virus-triggered activation of IRF3 and NF-κB, as well as IFN-β induction, whereas knockdown of TRIM4 had opposite effects. Mechanistically, TRIM4 associates with RIG-I and targets it for K63-linked polyubiquitination. Our findings demonstrate that TRIM4 is an important regulator of the virus-induced IFN induction pathways by mediating RIG-I for K63-linked ubiquitination.
-
Eight common genetic variants associated with serum DHEAS levels suggest a key role in ageing mechanisms. Zhai G, et al. PLoS Genet, 2011 Apr. PMID 21533175, Free PMC Article
tisdag 13 februari 2018
TRIM3 kuuluu p53 säätelyverkostoon. BERP- myös aivoissa
TRIM3 osalta löytyy merkitseviä kliinisiä tietoja
TRIM3 kuuluu ryhmään TRIM-NHL.
https://www.ncbi.nlm.nih.gov/pubmed/?term=TRIM3
Kaikki elimet joissa sitä ilmenee on kuvattu pylväin, eniten suolistossa, vähiten haimassa.
TRIM3 kuuluu ryhmään TRIM-NHL.
https://www.ncbi.nlm.nih.gov/pubmed/?term=TRIM3
Kaikki elimet joissa sitä ilmenee on kuvattu pylväin, eniten suolistossa, vähiten haimassa.
- BERP; HAC1; RNF22; RNF97
- Summary
- The protein encoded by this gene is a member of the tripartite motif (TRIM) family, also called the 'RING-B-box-coiled-coil' (RBCC) subgroup of RING finger proteins. The TRIM motif includes three zinc-binding domains, a RING, a B-box type 1 and a B-box type 2, and a coiled-coil region. This protein localizes to cytoplasmic filaments. It is similar to a rat protein which is a specific partner for the tail domain of myosin V, a class of myosins which are involved in the targeted transport of organelles. The rat protein can also interact with alpha-actinin-4. Thus it is suggested that this human protein may play a role in myosin V-mediated cargo transport. Alternatively spliced transcript variants encoding the same isoform have been identified. [provided by RefSeq, Jul 2008]
- Expression
- Ubiquitous expression in duodenum (RPKM 6.6), small intestine (RPKM 6.1) and 25 other tissues See more
- The tripartite motif-containing protein 3 on the proliferation and cytokine secretion of rheumatoid arthritis fibroblast-like synoviocytes. Wang M, et al. Mol Med Rep, 2017 Apr. PMID 28259936, Free PMC Article
- Potential role of TRIM3 as a novel tumour suppressor in colorectal cancer (CRC) development. Piao MY, et al. Scand J Gastroenterol, 2016. PMID 26691157
- Decreased expression of TRIM3 is associated with poor prognosis in patients with primary hepatocellular carcinoma. Chao J, et al. Med Oncol, 2014 Aug. PMID 24994609
- Human Brat ortholog TRIM3 is a tumor suppressor that regulates asymmetric cell division in glioblastoma. Chen G, et al. Cancer Res, 2014 Aug 15. PMID 24947043, Free PMC Article
GeneRIFs: Gene References Into FunctionsWhat's a GeneRIF?
Nivelreumassa TRIM 3 omaa antiproliferatiivisen tehtävän. JosTRIM3 esiintymä vahvennettaan yliexpressioon, p53 ja p21 säätyvät ylös. TNFalfa, Il1b, IL6 ja p38 säätyvät alas, samoin sykliini D1 ja PCNA, jotka ovat solusykliproliferaation edistäjiä. Solusyklin negatiivisia säätelijöitä ovat p53ja p21.
TRIM3 sitoutuu nukleaariseen importiinikompleksiin ja pitää yllä kantasolun tasapainoa; tuumorikasvua se vaimentaa säätelemällä NICD , aktiivin NOTC1:n tumaankuljetusta - syöpäsolupuolestaan häiritsee kantasolun uudistumisen ja multipotenssin solun erilaistumisen sekä asymmetrisen soluerilaistumisen keskeistä tasapainoa.
Cancer stem cells exert enormous influence on neoplastic behavior, in part by governing asymmetric cell division and the balance between self-renewal and multipotent differentiation. Growth is favored by deregulated stem cell division, which enhances the self-renewing population and diminishes the differentiation program......tripartite motif-containing protein 3 (TRIM3), similarly suppressed NOTCH1 signaling and markedly attenuated the stem cell component. We also found that TRIM3 suppressed nuclear transport of active NOTCH1 (NICD) in glioblastoma and demonstrated that these effects are mediated by direct binding of TRIM3 to the Importin complex. Together, our results support a novel role for Brat/TRIM3 in maintaining stem cell equilibrium and suppressing tumor growth by regulating NICD nuclear transport.
TRIM3 stabiloi p53:n ja siten voisi vaikuttaa colon tuumorin suppressiota mutta on tutkituissa colonkarsinomasoluissa alassäätyneenä.
TRIM kuuluu NHL-domeenin omaaviin TRIM- perheen jäseniin.
Tutkimusten johtopäätös on, että TRIM3 on maksasyövässä alassäätyneenä. Marköörinä sen alassäätymä on huonon prognoosin merkki. TRIM3 on mahdollinen terapeuttinen kohde hepatosellulaarisessa syövässä.- In conclusion, this study suggests that TRIM3
may play a significant role in HCC progression and acts as a valuable
prognostic marker and potential therapeutic target for HCC.
Cancer stem cells, capable of self-renewal and multipotent differentiation, influence tumor behavior through a complex balance of symmetric and asymmetric cell divisions. Mechanisms regulating the dynamics of stem cells and their progeny in human cancer are poorly understood.asymmetrically rather than symmetrically.
.... We conclude that TRIM3 acts as a tumor suppressor in GBM by restoring asymmetric cell division.
TRIM-perheen merkitys syöpäbiologiassa on kasvamaan päin. Tämä TRIM3 omaa NHL-domaanin ja sen kautta saattaa olla nisäkästuumoreitten vaimentaja. Jos TRIM3 ilmenemää vähennetään koehiirissä, niissä kiihtyy PDGF:n aiheuttama glioomakasvu. TRIM3 voi sitoutua CDK1-estäjään p21 ja kerryttää sitä, jolloin estyy CykliiniD1-CDK4 akkumulaatio . - reducing TRIM3 expression increased the incidence of and accelerated the development of platelet-derived growth factor -induced glioma in mice. Furthermore, TRIM3 can bind to the cdk inhibitor p21(WAF1/CIP1). Thus, we conclude that TRIM3 is a tumor suppressor mapping to chromosome 11p15.5 and that it might block tumor growth by sequestering p21 and preventing it from facilitating the accumulation of cyclin D1-cdk4.
-
p53 on keskeinen tekijä soluperäisessä stressivasteessa ja useimpien kasvannaisten vaimentajana. Kun on tunnistettu p53-signalointitien ainutlaatuisten molekyylien verkosto, voidaan saada selville tämän tärkeän transkriptiotekijän toimintoja . - Tässä artikkelissa kuvataan aivojen BERP tekijää (se on TRIM3). Sen geeni-ilmenemä on riippuvainen p53:sta, genomin suojelijasta. Niin ihmisessä kuin hiiressäkin. Poistogeenisillä tutkimuksilla todettiin , että aivoelektrisyydessä näkyi vaikutusta GABA-järjestelmän alueessa: postsynaptiset inhibitoriset virrat olivat alempiamplitudisia ja reseptorialayksikkömäärä väheni. Pääteltiin, että BERP (=aivojen ilmentämä RING finger proteiini, TRIM3) saattaa toimia posttranskriptionaalisella tasolla GABAreseptoreiden solunsisäisen liikehtimisen säätelyssä, sillä BERP- poisto ei kuitenkaan vähentänyt itse GABA- reseptorien mRNA:n määrää. Tutkijat pitävät TRIM3/BERP geenituotetta ainutlaatuisena p53:a säätelevänä tekijänä ja täten aivoissakin olisi osatehtävää p53:lle.
p53 is a central player in responses to cellular stresses and a major tumor suppressor. The identification of unique molecules within the p53 signaling network can reveal functions of this important transcription factor. Here, we show that brain-expressed RING finger protein (BERP) is a gene whose expression is up-regulated in a p53-dependent manner in human cells and in mice. We generated BERP-deficient mice by gene targeting and demonstrated that they exhibit increased resistance to pentylenetetrazol-induced seizures. Electrophysiological and biochemical studies of cultured cortical neurons of BERP-deficient mice showed a decrease in the amplitude of GABA(A) receptor (GABA(A)R)-mediated miniature inhibitory postsynaptic currents as well as reduced surface protein expression of GABA(A)Rs containing the gamma2-subunit. However, BERP deficiency did not decrease GABA(A)Rgamma2 mRNA levels, raising the possibility that BERP may act at a posttranscriptional level to regulate the intracellular trafficking of GABA(A)Rs. Our results indicate that BERP is a unique p53-regulated gene and suggest a role for p53 within the central nervous system
fredag 9 februari 2018
TRIM-NHL-perhe (TRIM2, TRIM3, TRIM32,TRIM71
http://www.cell.com/cms/attachment/603172/4766200/gr1.jpg
TRIM2 proteiini kuuluu TRIM-NHL-proteiinien perheeseen.
Tutut RBBCC domaanit toimivat E3 ubikitiiniligaasina ja NHL domaani (C-terminaalissa olevat 4-6 toistojaksot) tekevät interaktiota AGO-proteiinien kanssa.
Ihmisellä on useampia sellaisia TRIM-molekylejä joissa on C-terminaalinen NHL.
http://www.cell.com/cell/fulltext/S0092-8674%2809%2900206-2
TRIM-NHL on konservoitunut metazooilla ja näyttää säätelevän solun proliferaatiota ja kehitystä . Tässä artikkelissa niitä löydetään lisää ja ne ovat mikroRNA-funktion positiivisia säätelijöitä hiirillä.
The TRIM-NHL family of proteins is conserved among metazoans and has been shown to regulate cell proliferation and development. In this issue, Hammell et al. (2009) and Schwamborn et al. (2009) identify two members of this protein family, NHL-2 in worms and TRIM32 in mice, as positive regulators of microRNA function.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2374724/
Linkki tämän TRIM-NHL proteiinin merkityksestä ihmisellä-
Bardet-Biedl syndrome; Brat; LIN-41(TRIM71); Limb
girdle muscular dystrophy 2H; Mei-P26; NHL; Sarcotubular myopathy; TRIM;
TRIM32; TRIM71
{kind=link}
TRIM2 proteiini kuuluu TRIM-NHL-proteiinien perheeseen.
Tutut RBBCC domaanit toimivat E3 ubikitiiniligaasina ja NHL domaani (C-terminaalissa olevat 4-6 toistojaksot) tekevät interaktiota AGO-proteiinien kanssa.
Ihmisellä on useampia sellaisia TRIM-molekylejä joissa on C-terminaalinen NHL.
http://www.cell.com/cell/fulltext/S0092-8674%2809%2900206-2
TRIM-NHL on konservoitunut metazooilla ja näyttää säätelevän solun proliferaatiota ja kehitystä . Tässä artikkelissa niitä löydetään lisää ja ne ovat mikroRNA-funktion positiivisia säätelijöitä hiirillä.
The TRIM-NHL family of proteins is conserved among metazoans and has been shown to regulate cell proliferation and development. In this issue, Hammell et al. (2009) and Schwamborn et al. (2009) identify two members of this protein family, NHL-2 in worms and TRIM32 in mice, as positive regulators of microRNA function.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2374724/
Linkki tämän TRIM-NHL proteiinin merkityksestä ihmisellä-
TRIM-NHL-proteiinit ovat avainsäätelijöitä kehityksellisten transitioitten säätelyssä, esim. edistämässä erilaistumista estäen samalla solukasvua ja proliferaatiota kantasoluissa ja progeniittorisoluissa.
Tämän proteiinin epänormaaliuksilla on myös liittymänsä ihmistauteihin, erikoisesti sellaisiin tauteihin, jotka koskevat lihasten ja hermojen funktioita. Sen takia TRIM2-NHL kuten TRIM-NHL-proteiinit yleensä ovat mahdollisia terapeuttisen intervention kohteita. Tässä artikkelissaan tutkijat haluavat antaa systemaattisen ja laajan yhteenvedon useimmista tutkituista TRIM-NHL-proteiineista - erityisesti he antavat valaisevia esimerkkejä niistä, joissa on varmistettu yhteys rakenteellisiin piirteisiin, molekulaarisiin funktioihin ja biologisiin lopputuloksiin.
- TRIM-NHL proteins are key regulators of developmental transitions, for example promoting differentiation, while inhibiting cell growth and proliferation, in stem and progenitor cells. Abnormalities in these proteins have been also associated with human diseases, particularly affecting muscular and neuronal functions, making them potential targets for therapeutic intervention. The purpose of this review is to provide a systematic and comprehensive summary on the most studied TRIM-NHL proteins, highlighting examples where connections were established between structural features, molecular functions and biological outcomes.
- PMID:
- 26514622
- DOI:
- 10.1016/j.semcdb.2015.10.017
- [Indexed for MEDLINE]
2018-02-09 Koetan kirjoittaa näistä ryhmän jäsenistä erikseenkin niiden vastaavien kromosomien yhteydessä.
23:39
KUVA LIN-41 eli TRIM71 kaava:
NHL-toistojaksoista saa hyvän käsityksen esim tästä kaavakuvasta:
http://www.genetics.org/content/198/4/1535
KUVA LIN-41 eli TRIM71 kaava:
NHL-toistojaksoista saa hyvän käsityksen esim tästä kaavakuvasta:
TRIM1 , MID2 , Kr.Xq22.3
Geeniä ilmenee runsaiten kilpirauhasessa ja rasvakudoksessa ja 24 muussa kudoksessa,joista vähiten luuytimessä
Geenin nimiä ovat MID2, FXY2, RNF60, MRX101
http://www.cell.com/trends/biochemical-sciences/fulltext/S0968-0004%2817%2930002-6
Tässä yllä löytyy TRIM kartan alkua. PTEN tulee sijoittaa siihen , samoin p53 ja niiden suhde BRCA1 molekyyliin. Siitä on kuva netissä, muta jos sen otat tähän, koko sivu täyttyy kirjaimsita. HAkusana "PTEN BRCA1. Sitten voi liittää kartalla eri TRIM-molekyylit näihin tekijöihin.
PubMed linkki geenin kliinisistä merkityksistä:
Tämä X-kromosomaalinen TRIM1, MID2, on ubikitiinia konjugoiva E2-entsyymi ja assosioituu tuumorin etenemiseen ja se on tunnitettu BRCA1- geenin kanssa interaktiota tekeväksi. MID2 osuus kuitenkin rintasylövässä on ollut tuntematon ja tässä artikkelin tutkimuksessa selvitetään senosuutta, ilmenemää, prognostista arvoa rintasyövässä. MID2 mRNA ja proteiini olivat merktisevästi kohonnnet syöpäkudoksessa verrattunaa normaalin epiteelikudokseen. Arkistoiduista näytteistä 95.8% osoitti MID2 yli-ilmenemää. Lisäksi MID2 määrä kasvoi vakavan kliinisen vaiheen myötä. MID2 ilmenemä oli korkea vakavissa kliinisissä asteissa ja nousi T, N, M asteiden myötä. Katsotaan MID2:n olevan itsenäinen prognostinen tekijä huonommasta seuraamuksesta koko kohortissa. Koeputkitutkimuksissa havaittin MID2geeni hiljennyksellä syöpäsoluproliferaation vähenemää. In vivo xenograftituumorikin osoitti kasvun blokeerautumsita. Tämän tutkimuksen mukaan MID2 on uusi prognostinen merkitsijä ja myös rintasyövässä interventiokohteeksi soveltuva.
Front
Med. 2016 Mar;10(1):41-51. doi: 10.1007/s11684-016-0429-z. Epub
2016 Jan 20.
Midline2 is overexpressed and a prognostic indicator in human
breast cancer and promotes breast cancer cell proliferation in vitro
and in vivo.Wang L1, Wu J2, Yuan J2, Zhu X3,2, Wu H1, Li M4,5.
Midline2 (MID2) is an ubiquitin-conjugating E2 enzyme linked to tumor progression and a novel interacting partner of breast cancer 1, early-onset (BRCA1). However, the role of MID2 in breast cancer remains unknown. This study investigated the expression, prognostic value, and role of MID2 in breast cancer. The expression of MID2 mRNA and protein was significantly upregulated in breast cancer tissue and established cell lines compared with that in normal breast epithelial cells and paired adjacent non-tumor tissue (P < 0.001). Immunohistochemical analysis demonstrated that MID2 was overexpressed in 272 of 284 (95.8%) paraffinembedded, archived breast cancer tissue. Moreover, MID2 expression increased with advanced clinical stage (P < 0.001). High MID2 expression was significantly associated with advanced clinical stages and T, N, and M staging (all P < 0.05). Univariate and multivariate analyses indicated that high MID2 expression was an independent prognostic factor for poor overall survival in the entire cohort (93.73 vs. 172.1 months; P < 0.001, logrank test) and in subgroups with stages Tis + I + II and III + IV. Furthermore, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide colony formation, and anchorage-independent growth ability assays were conducted. Results showed that siRNA silencing of MID2 expression significantly reduced MCF-7 and MDA-MB-231 cell proliferation in vitro and blocked the growth of MDA-MB-231 cell xenograft tumors in vivo (P < 0.05). This study indicated that MID2 may be a novel prognostic marker and interventional target in breast cancer.
KEYWORDS:
MID2; breast cancer; overall survival; proliferation; xenograft
Musitiin 9.2. 2018 , 11:25.
Päivän sana Samuel Bagsterin kirjasta Valaistu Päivä
Klo 10:03
Samuel Bagster muistuttaa 9.2. aamupäivän tekstissä kirjassa Daily Light Johanneksen näystä Patmoksella Välimeren saarella kaksi tuhatta vuotta sitten .
Samuel Bagster muistuttaa 9.2. aamupäivän tekstissä kirjassa Daily Light Johanneksen näystä Patmoksella Välimeren saarella kaksi tuhatta vuotta sitten .
-Revelation 7: 13
Then one of the
elders addressed me, saying:
-”Who are these,
clothed in white robes, and whence have they come?”
I said to him:
-”Sir, you know.”
And he said to me:
-”These are they
who have come out of the great tribulation; they have washed their
robes and made them white in the blood of the Lamb. Therefore are
they before the throne of God, and serve Him day and night within
His temple; and He who sits upon the throne will shelter them with
His presence. They shall hunger no more, neither thirst any more; the
sun shall not strike them, nor any scorching heat. For the Lamb in
the midst of the throne will be their shepherd, and he will guide
them to springs of living water; and God will wipe away every tear
from their eyes.”
TRIM8 osuus glioomassa tutkittu Italiassa ensi kerran
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4468980/
BMC Cancer. 2015; 15: 470.
Published online 2015 Jun 16. doi: 10.1186/s12885-015-1449-9
PMCID: PMC4468980
TRIM 8 alassäätyminen gliomassa vaikuttaa soluproliferaatioon ja assosioituu potilaan elossapysymiseen. Ihmisen glioma on heterogeeninen ryhmä primäärejä maligneja aivotuumoreita, jonka molekulaarinen patogeneesi ei ole vielä selvinnyt. On tehty paljon tutkimusta, jotta löydetään gliomaan assosioituvia geenejä. Tässä artikkelissa selvitetään, miten TRIM8 geeni vaikuttaa glioomassa. Tutkijat osoittivat että TRIM8 ilmenemä korreloi epäsuotuisaan kliiniseen taudinkulkuun glioomapotilailla.
Jos palautettiin TRIM8 ilmenemä, indusoitui merkitsevä alenema klonogeenisessa U87MG soluviljelmässä ja potilaan glioblastoma soluissa. miR-17 voi kohdistua suoraan TIM8 3´UTR:ään ja posttranskriptonaalisesti vaimentaa TIM8- ilmenemää.
TRIM8 downregulation in glioma affects cell proliferation and it is associated with patients survival
Human gliomas are a heterogeneous group of primary malignant brain tumors whose molecular pathogenesis is not yet solved. In this regard, a major research effort has been directed at identifying novel specific glioma-associated genes. Here, we investigated the effect of TRIM8 gene in glioma.
We showed that TRIM8 expression correlates with unfavorable clinical outcome in glioma patients. We found that a restored TRIM8
expression induced a significant reduction of clonogenic potential in
U87MG and patient’s glioblastoma cells. Finally we provide experimental
evidences showing that miR-17 directly targets the 3′ UTR of TRIM8 and post-transcriptionally represses the expression of TRIM8.
http://www.mdpi.com/genes/genes-08-00354/article_deploy/html/images/genes-08-00354-g003.png
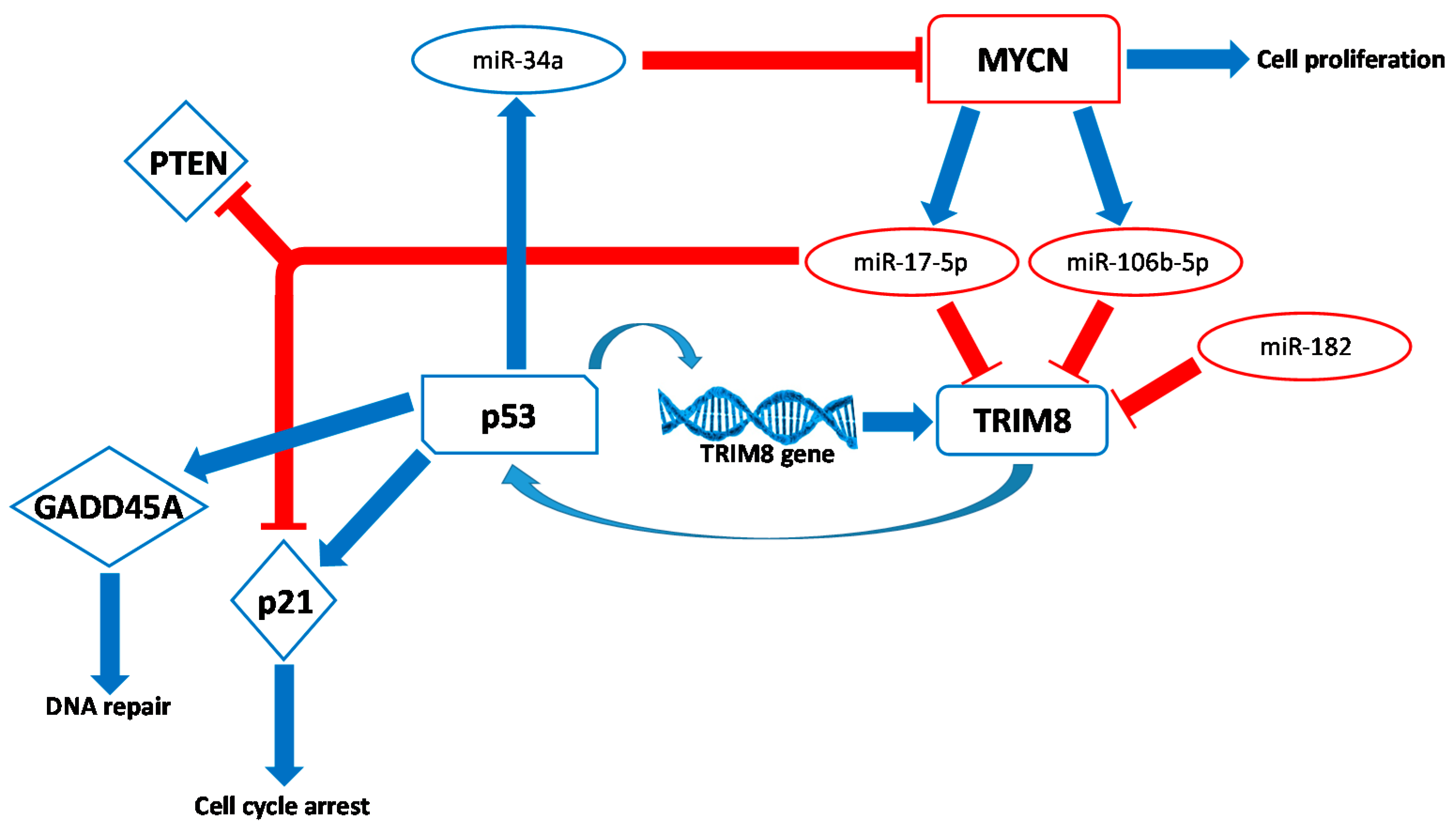{kind=link}
Muistiin 9.2. 2018 01:17
torsdag 8 februari 2018
TRIM24:n ja p53:n keskinäinen suhde DNA vaurioituneessa solussa
Tif-1alpha
(=TRIM24) ,kr.7q32-q34. Muita nimiä:RNF82, TIF1A,PTC6.välittää
transkriptiokontrollia, sijaitsee nukleaarisestikin ja vaikuttaa
hormonaalisiin reseptoreihin. Esiintyy runsaiten
hormonaalisissa elimissä. Vaikuttaa hormonaalisten elinten syöpien
etiologiassa mm. Säätelee negatiivisella feedbackillä p53.
Tästä viimeksimainitusta löydän kuvan: Se kertoo p53 (genomin suojelijan) ja TRIM24:n välisestä suhteesta. TRIM24 on tavallaan saman tapainen kuin MDM2 vaikutukseltaan.
TRIM24 kohdituu mieluiten sellaiseen p53 molekyliin joka on fosforyloitunut ja saataa sen ubikityloiduksi ja sitä tietä silppuroiduksi. Muta jos solua kohtaa esim IR-säteily ja DNA-vaurio, aktivoituu ATM-kinaasi ja fosforyloi p53:n ja paralleelisti se fosforyloio myös TRIM24:n. P53 aktivoituna siirtyy p53responsiiviseen elementiin TRIM24 geenissä ja alkaa tuotaa lisää TRIM24 molekyyliä. Silläaikaa fosforyloitunut TRIM24 on ehtinyt hajota ja autodestruktiivisesti ubikitinoituna kulkenut silppuriin. Uusi muodostunut TRIM24 ottaa sitten ubikitinoitavaksi ensisijaisesti fosforyloituneita p53-molekyyolejä. (ATMkinaasi estää samalla TRIM24:ää ubikitinoimasta fosforyloimatonta p53 molekyyliä). joten tässä nähdään negatiivinen feedback, jossa ei muodostunlisää p53:a.
http://mcb.asm.org/content/34/14/2695/F8.expansion.html

Musitiin 22:54 8.2. 2018
Tästä viimeksimainitusta löydän kuvan: Se kertoo p53 (genomin suojelijan) ja TRIM24:n välisestä suhteesta. TRIM24 on tavallaan saman tapainen kuin MDM2 vaikutukseltaan.
TRIM24 kohdituu mieluiten sellaiseen p53 molekyliin joka on fosforyloitunut ja saataa sen ubikityloiduksi ja sitä tietä silppuroiduksi. Muta jos solua kohtaa esim IR-säteily ja DNA-vaurio, aktivoituu ATM-kinaasi ja fosforyloi p53:n ja paralleelisti se fosforyloio myös TRIM24:n. P53 aktivoituna siirtyy p53responsiiviseen elementiin TRIM24 geenissä ja alkaa tuotaa lisää TRIM24 molekyyliä. Silläaikaa fosforyloitunut TRIM24 on ehtinyt hajota ja autodestruktiivisesti ubikitinoituna kulkenut silppuriin. Uusi muodostunut TRIM24 ottaa sitten ubikitinoitavaksi ensisijaisesti fosforyloituneita p53-molekyyolejä. (ATMkinaasi estää samalla TRIM24:ää ubikitinoimasta fosforyloimatonta p53 molekyyliä). joten tässä nähdään negatiivinen feedback, jossa ei muodostunlisää p53:a.
http://mcb.asm.org/content/34/14/2695/F8.expansion.html
Musitiin 22:54 8.2. 2018
Hematopoieettinen linja ja sen TRIM proteiinit
Siirrän tähän muistiin hematopoieettiseen linjaan vaikuttavia Trimejä
"(Sitaatti edellämainitusta artikkelista (TRIM35, HLS5)
The J2E erythroleukemic cell line has on rare occasions developed the features of monocytic cells.
Subtractive hybridization was used in an attempt to identify genes that were up-regulated during this erythroid to myeloid transition.
We report here on the isolation of hemopoietic lineage switch 5 (Hls5), a gene expressed by the monocytoid variant cells, but not the parental J2E cells.
Hls5 is a novel member of the RBCC (Ring finger, B box, coiled-coil)
family of genes,
which includes
Pml (TRIM19),kr.15q24.1. Muita nimiä: MYL, RNF71,PML (promyelosyyttileukemia) SumoE3 aktiivisuutta. Katso erikseen "TRIM19 ja verisyöpä "
Herf1 (=TRIM10), kr.6p21.3. (Alaluokka C IV PRY/SPRY) Muita nimiä: RNF9, Hematopoieettinen RING finger 1 RFB30. Tärkeä erytsosyyteille. Luuytimessä, munuaisessa.
Tif-1alpha (=TRIM24) ,kr.7q32-q34. Muita nimiä:RNF82, TIF1A,PTC6.välittää transkriptiokontrollia, sijaitsee nukleaarisestikin ja vaikuttaa hormonaalisiin reseptoreihin. Esiintyy runsaiten hormonaalisissaelimissä. Vaikuttaa hormonaalisten elinten syöpien etiologiassa mm. Säätelee negatiivisella feedbackillä p53. http://mcb.asm.org/content/34/14/2695/F8.expansion.html
Kts. erikseen "TRIM24 ja verisyöpä"
and
Rfp (TRIM27). kr.6p21. Muita nimiä:ZNFB7.Sijaitsee tumamatriisissa. E3-Ubikitiiniligaasiaktiivisuutta ja välittää PIK3C2B ubikitinoitumista ja estää sen aktiivisuutta. estää CD4Tsolujen aktivoitumista. Voi sitoutua dsDNA:han Esiintyy kilpirauhasessa, pernassa. Atyyppinen PTEN polyubikitylaatiovaara ja useita syöpiä, TRIM27 modifioi syöpäinsidenssiä. Myös PD taudissa TRIM27 on ylössäätynyt.
TRIM15 (Kr,6p21.33) RNF95, ZNF178, ( C IV) Fokaaliadheesiokomponentti
TRIM15 (Kr.6p21.33), RNF93, ZNF178, ZNFB7(Alaryhmä C IV PRY/SPRY)
TRIM15 geeni näyttää ilmenevän suolistoalueella sekä eritykseen osallistuvissa elimissä kuten maksa-sappirakko ja munuainen. Rakenne on alaluokan CIV mukainen: RING domaani, B-Box 1 ja B-Box2 , CC ja C-terminaalissa on PRY/SPRY.-
Also known as RNF93; ZNFB7; ZNF178 Summary The protein encoded by this gene is a member of the tripartite motif (TRIM) family. The TRIM motif includes three zinc-binding domains, a RING, a B-box type 1 and a B-box type 2, and a coiled-coil region. The protein localizes to the cytoplasm. Alternatively spliced transcript variants have been described, but their biological validity has not been determined. [provided by RefSeq, Jul 2008] Expression Biased expression in duodenum (RPKM 12.6), small intestine (RPKM 9.2) and 6 other tissues See more
Related articles in PubMed
-
A transcription map of the major histocompatibility complex (MHC) class I region. Gruen JR, et al. Genomics, 1996 Aug 15. PMID 8812418
-
TRIM15 is a focal adhesion protein that regulates focal adhesion disassembly. Uchil PD, et al. J Cell Sci, 2014 Sep 15. PMID 25015296, Free PMC Article
TRIM15 vuorovaikuttaa paxilliiniin ja pystyy täten negatiivisesti säätelemään solujen leviämistä ja migraatiota.
(Suomennosta:) Fokaalit adheesiot ovat makromolekulaarisia komplekseja, jotka liittävät aktiini-solutukirunkoa solun ulkoiseen matriisiin. Solun migroitumiselle on kriittisesti olennaista näiden fokaaliadheesioiden dynaaminen aineenvaihdunnallinen vaihtelu. Paxilliini on moniadaptoriproteiini, jolla on tärkeä osa fokaaliadheesiodynamiikan säätelyssä. Tässä artikkelissa tutkijat kertovat tunnistaneensa TRIM15- proteiinin sellaiseksi tekijäksi, joka asettuu paxilliinin kanssa vuorovaikutukseen ja on komponenttina fokaaliadheesioissa. TRIM-proteiinit saavat nimensä rakenteensa kolmiosaisuudesta. TRIM15 -proteiinin yksi rakenneosa on helikaalinen (C-C, coiled coil) domeeni ja sen domeenin välityksellä proteiini TRIM15 tekee interaktion paxilliinin N-terminaalisen LD2-motiiviin näissä fokaalisissa liittymissä- (myosiini-II:sta riippumattomalla tavalla). Siinä TRIM15 poikkeaa muista fokaaliadheesiokomponenteista, että se on stabiili ja sen liikkuvuus on rajoittunutta, koska se pystyy muodostamaan oligomeerejä. Jos TRIM15 puuttuu solusta, ne solut ovat huonoja migroitumaan ja niiden fokaaliadheesioiden muodostumien on alentunutta. Johtopäätös: TRIM15:n solufunktiona on toimia fokaaliadheesion vaihtuvuuden ja solu migraation säätelykomponenttina
GeneRIFs: Gene References Into FunctionsWhat's a GeneRIF?
(
Suomennosta) Tässä artikkelin työssä tutkijat ovat osoittaneet,
että TRIM15 asettuu fokaaliadheesioihin dimerisoitumalla ja
merkitsevästi vaimentaa solumigraatiota. B-box2 ja PRY-domeenit
ovat essentiellit , jotta TRIM15 pystyy lokalisoitumaan
fokaaliadheesioihin ja estämään solujen migroitumisen. Havaittiin
myös että TRIM15 tekee interaktioita
(integriiniadhesomitutkimuksessa) useaan proteiiniin, joista
mainitaan koroniini 1B, kortaktiini, filamiinin sitoutuva
LIMproteiini1 ja vasodilataattorilla stimuloituva fosfoproteiini ,
jotka osallistuvat aktiini-sytoskeleton dynamiikkaan. TRIM15 ilmenemä
on kudoksiin rajoittuva ja paksunsuolen syövässä se on
alassäätynyt. TRIM15 esiintymispitoisuudet assosioituivat
kolonkarsinoomasolujen migroitumiseen ja tuumorin kasvuun. Tämä
tutkimus antaa uutta oivallusta TRIM15: n osuuteen
integriiniadhesomin lisäkomponenttina, solumigraation säätelijänä
viitaten siihen, että TRIM15 saattaa toimia paksunsuolituumorin
suppressorina.
-
The tripartite motif containing (TRIM) proteins are a large family of proteins that have been implicated in many biological processes including cell differentiation, apoptosis, transcriptional regulation, and signaling pathways. Here, we show that TRIM15 co-localized to focal adhesions through homo-dimerization and significantly suppressed cell migration. Domain mapping analysis indicated that B-box2 and PRY domains were essential for TRIM15 localization to focal adhesions and inhibition of cell migration. Our protein-protein interaction screen of TRIM15 with the integrin adhesome identified several TRIM15 interacting proteins including coronin 1B, cortactin, filamin binding LIM protein1, and vasodilator-stimulated phosphoprotein, which are involved in actin cytoskeleton dynamics. TRIM15 expression was tissue-restricted and downregulated in colon cancer. Level of TRIM15 expression was associated with colon cancer cell migration, as well as both in vitro and in vivo tumor growth. These data provide novel insights into the role of TRIM15 as an additional component of the integrin adhesome, regulating cell migration, and suggest that TRIM15 may function as a tumor suppressor of colon cancer.
Ilmeisesti TRIM15
osallistuu paxilliinin säätelyyn ja paxilliinin ylimääräinen
ilmeneminen on tumorogeenista:
Paxilliini on tärkeä
adaptorimolekyyli. Katson sen rakennetta ja funktion ulottuvuutta.
Sillä voi olla osuutta moniin signaaliteihin. Siinä on monia
motiiveja, joiden välityksellä se tekee
proteiini-proteiiniliittymiä. LD- motiivin kautta se tekee liittymän
TRIM15 proteiiniin. Ja näitten motiiviensa välityksellä se
toimii monen sytoskeletaalisen proteiinin alustana ja ne taas
rekrytoivat lisää proteiineja tähän fokaalisen
adheesiokompleksiin. Paxilliini ilmeisesti osaltaan koordinoi
signaaleita , joista sitten aiheutuu lopulta solujen leviäminen ja
liikkuvuus. Paxilliini esiintyy laajemmin kudoksissa kuin TRIM15, (
TRIM15 vaikutus on rajoittunut ja on paikallinen rajoittuen lähinnä
suolistoon, munuaiseen, sappirakkoon ja maksaan)
-
https://www.ncbi.nlm.nih.gov/pubmed/11607845 Paxillin is a focal adhesion-associated, phosphotyrosine-containing protein that may play a role in several signaling pathways. Paxillin contains a number of motifs that mediate protein-protein interactions, including LD motifs, LIM domains, an SH3 domain-binding site and SH2 domain-binding sites. These motifs serve as docking sites for cytoskeletal proteins, tyrosine kinases, serine/threonine kinases, GTPase activating proteins and other adaptor proteins that recruit additional enzymes into complex with paxillin. Thus paxillin itself serves as a docking protein to recruit signaling molecules to a specific cellular compartment, the focal adhesions, and/or to recruit specific combinations of signaling molecules into a complex to coordinate downstream signaling. The biological function of paxillin coordinated signaling is likely to regulate cell spreading and motility. Clinical significanceof paxillin. Paxillin has been shown to have a clinically-significant role in patients with several cancer types. Enhanced expression of paxillin has been detected in premalignant areas of hyperplasia, squamous metaplasia and goblet cell metaplasia, as well as dysplastic lesions and carcinoma in high-risk patients with lung adenocarcinoma.[24] Mutations in PXN have been associated with enhanced tumor growth, cell proliferation, and invasion in lung cancer tissues.[25] During tumor transformation, a consistent finding is that paxillin protein is recruited and phosphorylated.[26] Paxillin plays a role in the MET tyrosine kinase signaling pathway, which is upregulated in many cancers.[27]
Päivitys 4.4. 2018
Prenumerera på:
Kommentarer (Atom)